
Download
Score = Total score 2 × number of criteria applicable × 100 %
REVIEW ARTICLE
Aerotolerance of Campylobacter spp. in food chain: a meta-analysis on the prevalence and a systematic review on the persistence, genetic relatedness, and risk to humans
Kah Hui Chong1, Nor Ainy Mahyudin2,3, Hanan Hasan1, Chia Yeung New4, Nur Shahera Mohammad Sabri1, Shan Jiang1, Kousalya Padmanabhan1, Nor Azmiraah Abd Jabar5, Wanyu Pan5, Son Radu1, Mahmud Ab Rashid Nor-Khaizura1,5*
1Department of Food Science, Faculty of Food Science and Technology, Universiti Putra Malaysia, 43400, UPM Serdang, Selangor, Malaysia;
2Department of Food Service and Management, Faculty of Food Science and Technology, Universiti Putra Malaysia, 43400, UPM Serdang, Selangor, Malaysia;
3Halal Products Research Institute, Universiti Putra Malaysia, 43400, UPM Serdang, Selangor, Malaysia;
4Go Plus Services Sdn. Bhd., 97A, Jalan BP 6/3, Bandar Bukit Puchong, 47120, Puchong, Selangor, Malaysia;
5Laboratory of Food Safety and Food Integrity, Institute of Tropical Agriculture and Food Security, Universiti Putra Malaysia, 43400, UPM Serdang, Selangor, Malaysia
Abstract
Campylobacter spp. are the bacterial pathogens that cause gastrointestinal illness in humans, and were once asserted to be a microaerophile and unable to survive aerobically. In recent decades, aerotolerant Campylobacter spp. have been reported in the food chain. This review aims to obtain information on the aerotolerant Campylobacter spp., highlighting the prevalence, persistence, genetic relatedness, and risk to humans. After screening, the findings of 39 articles (12 articles for meta-analysis) were synthesized. High pooled prevalence of aerotolerant Campylobacter spp. (75.9%) from meta-analysis indicated the elevated risk of human campylobacteriosis. Campylobacter spp. with a higher aerotolerance level survived better against environmental stresses, including atmospheric, chemical antimicrobial agents, temperature, and osmotic conditions. The situation worsens with reported aerobic growth of Campylobacter spp. The sequence type (ST) of Campylobacter spp. had a statistically significant (P < 0.001) influence on aerotolerance level, suggesting that future studies would be able to postulate the aerotolerance level based on the determined ST. Aerotolerant Campylobacter spp. might pose an increased risk to humans because of their prevalence and persistence in a food chain, possible greater antibiotic resistance, and higher frequency of virulence genes. This underscores the potentially elevated medical burden of this pathogen.
Key words: Campylobacter jejuni, Campylobacter coli, meta-analysis, survival, food safety risk, antibiotic resistance
*Corresponding Author: Mahmud Ab Rashid Nor-Khaizura, Department of Food Science, Faculty of Food Science and Technology, Universiti Putra Malaysia, 43400, UPM Serdang, Selangor, Malaysia. Email: [email protected]
Academic Editor: Mehran Moradi, PhD, Department of Food Hygiene and Quality Control, Faculty of Veterinary Medicine, Urmia University, Urmia, Iran
Received: 26 June 2025; Accepted: 6 January 2026; Published: 16 April 2026
© 2026 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Since 2005, Campylobacter spp. have been reported as the most common gastrointestinal bacterial pathogen in the European Union (EU), accounting for 148,181 incidences of human campylobacteriosis in 2023 (European Food Safety Authority [EFSA] and European Centre for Disease Prevention and Control [ECDC], 2024). Campylobacter spp. affect more than 1.5 million people annually in the United States (Centers for Disease Control and Prevention [CDC], 2024). Campylobacter jejuni (C. jejuni) contributes around 90% of the human campylobacteriosis cases, while Campylobacter coli (C. coli) accounts for approximately 10% (EFSA and ECDC, 2024; Ministry of Health Singapore, 2025). Abdominal cramping, fever, and diarrhea with or without bloody stool are the clinical symptoms of campylobacteriosis, and commonly, these are indistinguishable from other bacterial gastrointestinal illnesses (Baek et al., 2024; Iversen et al., 2024; Kelly and Hodges, 2024). Symptoms may persist for 3–10 days, with an incubation period of 1–11 days (Myintzaw et al., 2023). Campylobacteriosis leads to sodium malabsorption syndrome, the severity of which depends on the type of strain and patient’s immune status (Bücker et al., 2018; Imbrea et al., 2024). However, on some occasions, post-infectious sequelae may develop, such as Miller Fisher syndrome (MFS), Guillain–Barré syndrome (GBS), reactive arthritis, and intestinal tract chronic inflammatory conditions, with a latent period of weeks or more (Backert et al., 2017; Heimesaat et al., 2023; Keithlin et al., 2014).
In an oxygen requirement and tolerant study conducted by Kaakoush et al. (2007), C. jejuni was classified as an obligate microaerophile. It was purported that the microaerophilic propensity of Campylobacter spp. is adapted to low oxygen concentration in the avian gut (Park, 2002). Campylobacter spp. grow in microaerobic conditions, with oxygen concentration ranging from 2.5% to 15%, compared to the atmospheric oxygen level of 21% (Haines et al., 2011; Kaakoush et al., 2007; Lynch et al., 2011). As Campylobacter spp. are transmitted from animals to humans zoonotically through food, the survivability of the pathogen outside host animal against environmental stress is crucial for its transmission to humans (Begley and Hill, 2015). Aerobic tolerance (aerotolerance) is a key survival mechanism for Campylobacter spp. in the food industry, where the oxygen level is high for the survival of pathogens (ca. 21%) (Kim et al., 2019). Aberrant aerotolerant Campylobacter spp. have been increasingly reported in various meats and viscera, including chicken meat, duck meat, turkey, pork, chicken livers, chicken gizzards, and beef livers (Guk et al., 2021; Karki et al., 2018; Oh et al., 2015a; Song et al., 2020). Aerotolerance levels of Campylobacter spp. can be classified into aero-sensitive (OS, did not survive after 12 h of aerobic incubation), aerotolerant (AT, survived after 12 h of aerobic incubation), and hyperaerotolerant (HAT, survived after 24 h of aerobic incubation) (Oh et al., 2015a). This review aims to compile information on AT Campylobacter spp. in terms of prevalence, environmental persistence, genetic relatedness, and the potential risks for humans.
Methodology
Search structure and strings applied
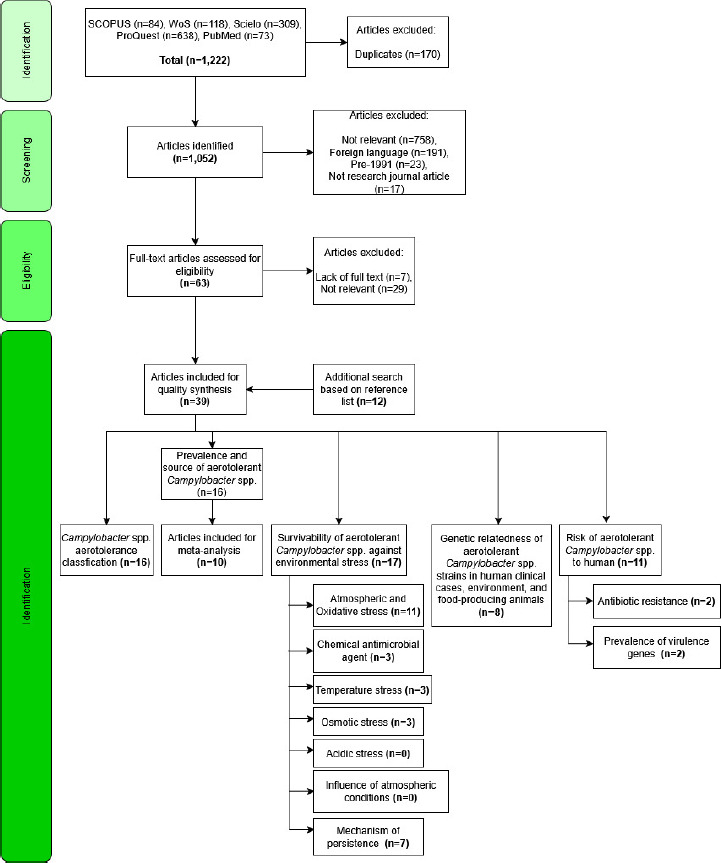
The search steps were completed using the Preferred Reporting Items for Systematic Reviews and Meta-analyses (PRISMA) guidelines (Page et al., 2021). The search was conducted in five electronic bibliographic databases, such as Scopus, Web of Science, PubMed, Scielo, and ProQuest, to increase the chance of identifying all potentially relevant articles. The date of the search was 10 January 2025. The search string used in Scopus, Web of Science, PubMed, and ProQuest was aerotoleran* AND campylobacter, while the search string used in Scielo was (Aerotolerance) OR (Aerotolerant) AND (Campylobacter). Figure 1 depicts the scheme of the study design describing the systematic review and meta-analysis based on PRISMA. The systematic review and meta-analysis protocol is not registered in any platform.
Figure 1 The scheme of study design describes the systematic review based on the PRISMA guidelines.
Eligibility criteria
Inclusion criteria
The inclusion criteria considered articles linking Campylobacter spp. to aerotolerance. No limitations were imposed on geographic location and sample size. Only peer-reviewed research articles published in English were included.
Exclusion criteria
The exclusion criteria barred all articles published before 1991, as studies prior to this year might not have accurately reported the true Campylobacter spp. (Vandamme et al., 1991, 1992). Further, articles that lack full text were also excluded. In addition, despite aerotolerance closely pertaining to oxidative stress, studies related to oxidative stress without descriptions on the aerotolerance phenotype were outside the remit of this study. Moreover, articles that reported the prevalence of AT Campylobacter spp. were excluded when the sample size was less than 30 (Je et al., 2024).
Screening process
Records were imported into Rayyan, and duplicates were removed (Ouzzani et al., 2016). Articles were then screened by comparing the inclusion and exclusion criteria on both title and abstract. Next, full-text articles were assessed for relevance. After reviewing the bibliography of the articles included, additional eligible studies were included. Based on the content of the included articles, several subtopics that built the body of this manuscript were synthesized as shown in Figure 1.
Two co-authors screened the articles independently based on the aforementioned inclusion/exclusion criteria. The included and excluded articles were based on the agreement between the two co-authors. When the agreement was not met, the final decision was made by the third co-author.
Risk of Bias Assessment (quality assessment)
The quality assessment tool for in vitro studies (QUIN tool) was adapted from Sheth et al. 2024. The QUIN tool allows researchers to assess the quality and risk bias of the included studies. The QUIN tool in this study consists of eight applicable criteria, as shown in Table S1. Each criterion was scored as follows: two points when adequately specified, one point when inadequately specified, and zero point when not specified. The scoring guidelines grade the study into high risk of bias (<50%), medium risk of bias (50–70%), and low risk of bias (>70%), by the following formula:
Meta-analysis
A proportional meta-analysis on the prevalence of AT Campylobacter spp. isolates was conducted with the Comprehensive Meta-Analysis software version 3.7 (Biostat Inc., Englewood, NJ, USA) (Borenstein, 2022). The proportion of AT Campylobacter spp. in this meta-analysis comprised both HAT and AT isolates. Prevalence with a 95% confidence interval (95% CI) was quantified based on the total sample and the corresponding positive number. Forest plots were constructed to illustrate the prevalence of individual studies along with the pooled estimate within 95% CI. Random-effect model was employed to account for both sampling error (within-study variance) and statistical heterogeneity (between-study variance) (Kanters, 2022). Heterogeneity was assessed using Cochran’s Q statistic and I2 inconsistency index. The estimated pooled prevalence can be inflated if many of the included studies have no events and have relatively low sample sizes (Churchill et al., 2019). Thus, meta-analyses were limited to the studies with a minimum sample size of 30 (Je et al., 2024).
Meta-regression and a priori subgroup meta-analysis were conducted to assess the potential reasons for between-study heterogeneity. Prevalence estimates were compared with three potential moderator variables (subgroups): source of AT Campylobacter spp. isolates (chicken, duck, and humans), species of AT Campylobacter spp. (C. jejuni and C. coli), and aerotolerance level of Campylobacter spp. (HAT and AT). Subgroup meta-analysis was performed if more than two studies were obtained in each subgroup (Plishka et al., 2021). In the subgroup of the source of AT Campylobacter spp. isolates, organs, feces, and meat samples were regrouped and analyzed based on their respective animal sources. In addition to the above-mentioned moderators, the potential covariates, including the publication year and location, were assessed with meta-regression.
Publication bias was assessed with the funnel plot, Egger’s weighted regression test, and fail-safe N test (Egger et al., 1997; Rothstein et al., 2005). An asymmetric funnel plot provides visual judgment for publication bias (Borenstein et al., 2009). Egger’s test is a statistical test to discover funnel plot asymmetry (van Enst et al., 2014). The fail-safe N test computes the number of missing studies (N) required to incorporate meta-analysis before the P value becomes nonsignificant (Rosenthal, 1979). If the N > 5 k +10, where k is the number of studies incorporated in the meta-analysis, the robustness of the meta-analysis can be assured (Rosenthal, 1979).
Statistical analysis
An alluvial plot illustrated the overview of Campylobacter spp. isolates with different levels of aerotolerance from several sources studied against environmental stresses was generated using the Origin 2023b (OriginLab Corporation, MA, USA). Ordinal logistic regression was performed to investigate the relationship between Campylobacter spp. sequence type (ST) and aerotolerance level with the Minitab 19 statistical software (Minitab, LLC, PA, USA). Radial graphs of clonal complexes (CC) and ST of C. jejuni and C. coli with different aerotolerance levels were illustrated with the Flourish platform (flourish.studio/).
Results and Discussion
Characteristics of articles and risk of bias
In all, 1,222 articles were retrieved from SCOPUS, Web of Science, Scielo, ProQuest, and PubMed. After screening, 39 peer-reviewed articles were included in this review. In the screening of the title and abstract, 989 articles were excluded (758 studies were not relevant, 191 studies were in a foreign language, 23 studies were pre-1991, and 19 studies were not research journal articles). During full-text assessment, 7 articles were excluded due to the lack of full text, and 20 articles that were not relevant were eliminated. Next, 10 studies were included after an additional search based on the reference list. All studies included in the pooled analysis reported positive samples for AT Campylobacter spp.
The included studies were synthesized into several sub-topics: 16 studies for Section “Campylobacter spp. aerotolerance classification”, 16 studies for Section “Prevalence and source of aerotolerant Campylobacter spp.”, 17 studies for Section “Survivability of aerotolerant Campylobacter spp. against environmental stress”, 8 studies for Section “Genetic relatedness of aerotolerant Campylobacter spp. strains”, and 11 studies for Section “Risk of aerotolerant Campylobacter spp. to humans”, as demonstrated in Figure 1. As shown in Table S1, most studies were considered to have a low risk of bias (>70%), and only one study was identified with a medium risk of bias (68.75%).
Aerotolerant Campylobacter spp. were first reported in the 1970s (Hanna et al., 1983; Neill et al., 1979). The genus Arcobacter was first proposed in 1991 based on nucleic acid hybridization and other molecular studies of AT Campylobacter (Vandamme et al., 1991). Arcobacter species were distinguished from the true Campylobacter spp. by their capability to grow between 15°C and 30°C and their aerotolerance properties (Vandamme et al., 1991, 1992). From that, Campylobacter cryaerophila and Campylobacter butzleri were proposed to be classified under arcobacter (Vandamme et al., 1991, 1992).
Campylobacter spp. aerotolerance classification
In total, 16 studies classified Campylobacter spp. isolates based on the level of aerotolerance, while 14 studies investigated C. jejuni, and 5 studies investigated C. coli as shown in Table 1. The majority of studies referenced Oh et al. (2015a), probably because of its simplicity and clear-cut aerotolerance classification. From that, aerotolerance levels of C. jejuni were classified after aerobic incubation in Mueller Hinton (MH) broth with shaking at 200 revolutions per minute (rpm) at 42°C into OS (did not survive after 12 h), AT (survived after 12 h), and HAT (survived after 24 h) (Oh et al., 2015a). The term ‘intermediate aerotolerant’ was adopted by Pokhrel et al. (2023) for AT. In comparison, Lee et al. (2019) classified AT C. jejuni with survival after 120 h under 500 rpm aerobic shaking at 37°C. Furthermore, Jones et al. (1993) and Shagieva et al. (2021) classified C. jejuni as aerotolerant if colonies grew after aerobic incubation at 37°C or 42°C, respectively. Although Chynoweth et al. (1998) did not classify aerotolerance in their study, the ability of C. jejuni to grow aerobically on an agar plate was reported and was regarded as aerotolerant in this review.
Table 1 List of 16 studies that classified Campylobacter spp. based on aerotolerance.
| Study | Campylobacter spp. | Aerotolerant classification |
|---|---|---|
| Ortega-Sanz et al., 2024 | C. jejuni | Follow Oh et al. (2015a) |
| Pokhrel et al., 2023 | C. jejuni | Follow Oh et al. (2015a) |
| Guk et al., 2021 | C. coli | Follow Oh et al. (2015a) |
| Mouftah et al., 2021 | C. jejuni and C. coli | Follow Oh et al. (2015a) |
| Shagieva et al., 2021 | C. jejuni | Able to form colony under aerobic conditions |
| Jaakkonen et al., 2020 | C. jejuni | Follow Oh et al. (2015a) |
| Guk et al., 2019 | C. coli | Follow Oh et al. (2015a) |
| Karki et al., 2019 | C. jejuni and C. coli | Follow Oh et al. (2015a) |
| Kiatsomphob et al., 2019 | C. jejuni | Follow Oh et al. (2015a) |
| Kim et al., 2019 | C. jejuni | Follow Oh et al. (2015a) |
| Lee et al., 2019 | C. jejuni | 2 log10 CFU/mL reduction for 120 h as a threshold |
| Karki et al., 2018 | C. jejuni and C. coli | Follow Oh et al. (2015a) |
| Oh et al., 2018 | C. jejuni | Follow Oh et al. (2015a) |
| Oh et al., 2015a | C. jejuni | Classified into aero-sensitive (OS, did not survive after 12 h of aerobic incubation), aerotolerant (AT, survived after 12 h of aerobic incubation), and hyperaerotolerant (HAT,survived after 24 h of aerobic incubation), after aerobic incubation in MH broth with 200 rpm shaking at 42°C |
| Chynoweth et al., 1998 | C. jejuni | Capable of aerobic growth on a nutrient agar plate |
| Jones et al., 1993 | C. jejuni | Capable of aerobic growth on a blood agar plate |
Note: CFU/mL: Colony Forming Units per milliliter; MH: Mueller Hinton.
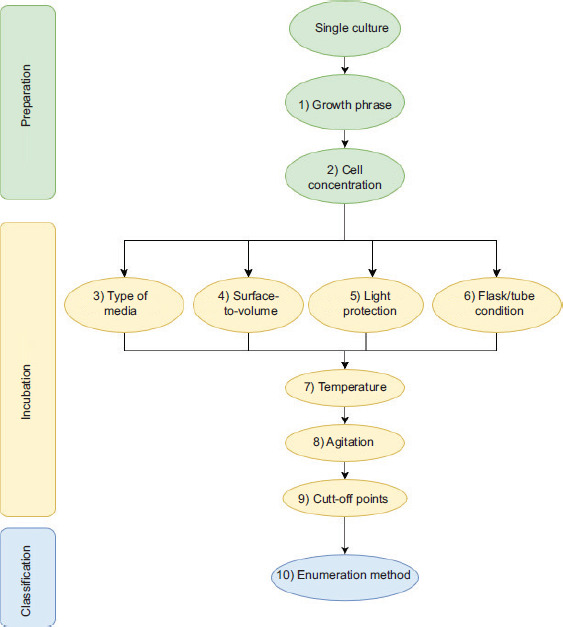
While the majority of the researchers adopted the method reported by Oh et al. (2015a), the protocol did not specify details, such as the surface-to-volume ratio of MH broth during aerobic incubation. Aerotolerance study in Campylobacter spp. is relatively new. Thus, a standardized method for determining the level of aerotolerance is crucial for inter-laboratory comparison, which may consider the following aspects: (1) growth phase of the inoculating culture (after certain hours of incubation); (2) cell concentration; (3) type of inoculating media; (4) aerobic stress of incubation, such as the surface-to-volume ratio of the media; (5) incubation flask/tube is protected from light/unprotected; (6) incubation flask/tube is tightened/cracked/loosened/left open; (7) incubation temperature; (8) degree and type of agitation during incubation; (9) cut-off points for aerobic incubation duration for aerotolerance level classification; and (10) enumeration method. The schematic diagram for the standardization protocol is shown in Figure S1.
Prevalence and source of aerotolerant Campylobacter spp.
Studies on the prevalence of AT Campylobacter spp. are limited, and the 16 published studies are summarized in Table S2. The prevalence of AT strains varies considerably between studies, ranging from 0% to 100% in C. jejuni and 41.9% to 100% in C. coli. Shagieva et al. (2021) reported an inconsistent AT profile of C. jejuni over three replicates. Thus, the three strains that consistently survived aerobically in all three replicates are labeled as AT in Table S2.
The Campylobacter spp. reported in studies stated in Table S2 are sorted by the source of the strains and presented in Table 2. Pond water and the outlet of a wastewater treatment plant from Shagieva et al. (2021) and udder cloth from Jaakkonen et al. (2020) were regrouped into the classification of environment, as shown in Table 2. Moreover, C. jejuni (697 strains) were characterized for aerotolerance level than C. coli (300 strains). Among them, chicken and pork were the most studied sources for aerotolerance in C. jejuni and C. coli, respectively. In C. jejuni, dairy product had the highest prevalence of HAT (69.5%), while duck had the highest prevalence of AT (62.2%). In C. coli, humans were found to have the highest prevalence of HAT (60%), and duck was reported to have the highest prevalence of AT (38.7%). In both C. jejuni and C. coli, AT (39.9% in C. jejuni and 35.2% in C. coli) and HAT (32.9% in C. jejuni and 44.3% in C. coli) were more prevalent than the OS counterparts. These estimates indicated that the AT strains of Campylobacter spp. could be an emerging phenomenon.
Table 2 Prevalence of Campylobacter spp. with different levels of aerotolerance from different sources as discovered in 16 studies.
| Source | C. jejuni | C. coli | ||||
|---|---|---|---|---|---|---|
| OS | AT | HAT | OS | AT | HAT | |
| Chicken | 38.8% (112/289) | 34.3% (99/289) | 27% (78/289) | 14.9% (11/74) | 37.8% (28/74) | 47.3% (35/74) |
| Beef | 20% (8/40) | 42.5% (17/40) | 37.5% (15/40) | 36.8% (7/19) | 31.6% (6/19) | 31.6% (6/19) |
| Dairy products | 8.7% (2/23) | 21.7% (5/23) | 69.5% (16/23) | 100% (2/2) | 0% (0/2) | 0% (0/2) |
| Duck | 13.3% (6/45) | 62.2% (28/45) | 24.4% (11/45) | 16.1% (10/62) | 38.7% (24/62) | 45.2% (28/62) |
| Humans | 10.3% (22/213) | 41.3% (88/213) | 48.4% (103/213) | 20% (3/15) | 20% (3/15) | 60% (9/15) |
| Environment | 61.4% (35/57) | 28.1% (16/57) | 10.5% (6/57) | 0 | 0 | 0 |
| Pork | 0 | 0 | 0 | 14.3% (72/126) | 28.6% (36/126) | 57.1% (18/126) |
| Turkey | 100% (5/5) | 0 | 0 | 0% (0/2) | 100% (2/2) | 0% (0/2) |
| N/A | 0% (0/25) | 100% (25/25) | 0% (0/25) | 0 | 0 | 0 |
| Total | 27.3% (190/697) | 39.9% (278/697) | 32.9% (229/697) | 20.5% (105/300) | 35.2% (99/300) | 44.3% (96/300) |
Notes: OS: aero-sensitive; AT: aerotolerant; HAT: hyperaerotolerant; N/A: not available.
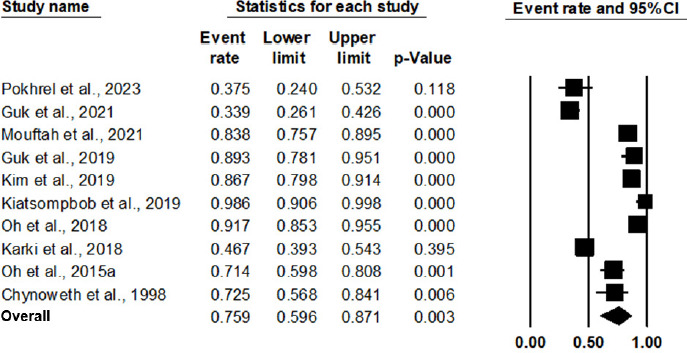
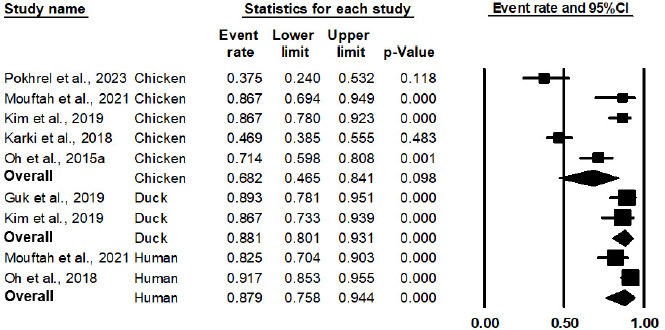
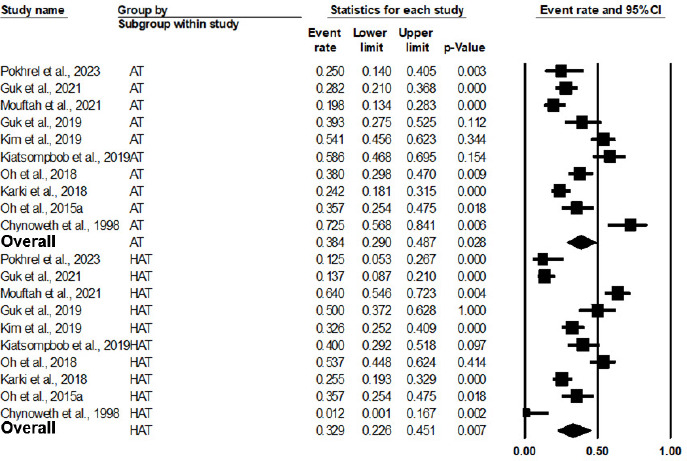
As shown in Figure 2 and Table 3, the pooled prevalence of AT Campylobacter spp. from 10 studies in a meta-analysis was 75.9% (95% CI: 59.6% to 87.1%, P = 0.003). The outputs of the subgroup analysis are shown in Figure 3 and Table 3; the pooled prevalence of AT Campylobacter spp. in chicken was 68.2% (95% CI: 46.5% to 84.1%, P = 0.098). As a comparison, the pooled prevalence of AT Campylobacter spp. in duck and humans was statistically significant (P < 0.001), with a pooled prevalence of 88.1% (95% CI: 80.1% to 93.1%) and 87.9% (95% CI: 75.8% to 94.4%), respectively. Based on meta-regression, as shown in Table 4, no statistically significant difference exists in the prevalence of AT Campylobacter spp. between chicken and duck, with P = 0.1226, and between chicken and humans, with P = 0.1165. Owing to the limitation of a 30-sample size in the respective studies, the source of AT Campylobacter spp., such as beef, dairy products, the environment, and turkey, were not included in the meta-analysis. This implies that further studies involving larger sample sizes from different sources are noteworthy for meta-analysis in the future.
Figure 2 Forest plot of the estimated prevalence of aerotolerant Campylobacter spp.
Figure 3 Forest plot of the estimated prevalence of aerotolerant Campylobacter spp. by subgroup of the source of isolates.
Table 3 Result of the overall meta-analysis and subgroup analysis on the prevalence of aerotolerant Campylobacter spp. based on three potential effect modifiers.
| Variables | Number of studies | Number of isolates | Number of positive isolates (HAT and AT) | Pooled prevalence and 95% confidence interval (CI) | Heterogeneity | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Pooled prevalence (%) | Lower limits (%) | Upper limits (%) | τ2 | Q value | I2 (%) | P value | ||||
| Overall | 10 | 934 | 654 | 75.9 | 59.6 | 87.1 | 1.364 | 177.897 | 94.941 | <0.001 |
| Source | ||||||||||
| Chicken | 5 | 360 | 230 | 68.2 | 46.5 | 84.1 | 0.947 | 50.411 | 92.065 | <0.001 |
| Duck | 2 | 101 | 89 | 88.1 | 80.1 | 93.1 | 0 | 0.163 | 0 | 0.687 |
| Humans | 2 | 178 | 158 | 87.9 | 75.8 | 94.4 | 0.254 | 3.207 | 68.819 | 0.073 |
| Species | ||||||||||
| C. jejuni | 8 | 642 | 476 | 73.7 | 50.1 | 88.7 | 1.634 | 130.478 | 94.635 | <0.001 |
| C. coli | 3 | 271 | 175 | 73.7 | 38.8 | 92.5 | 1.996 | 46.123 | 95.664 | <0.001 |
| Aerotolerance level | ||||||||||
| HAT | 10 | 924 | 323 | 32.9 | 22.6 | 45.1 | 0.578 | 100.198 | 91.018 | <0.001 |
| AT | 10 | 924 | 341 | 38.4 | 29.0 | 48.7 | 0.399 | 77.776 | 88.428 | <0.001 |
Notes: HAT: hyperaerotolerant; AT: aerotolerant.
τ2: tau-squared; I2: inconsistency index; Q: Cochran’s Q statistic.
Table 4 Meta-regression of the overall study and the subgroup.
| Q model | P value | Residual I2(%) | Residual τ2 | R2 (%) | |
|---|---|---|---|---|---|
| Overall | |||||
| Publication year | 3.89 | 0.5653 | 96.22 | 2.3112 | 0 |
| Location | 1.07 | 0.7839 | 96.13 | 1.9920 | 0 |
| Subgroup | |||||
| Source | 3.80 | 0.1497 | 88.84 | 0.7587 | 35.00 |
| Chicken* | – | – | – | – | – |
| Duck | – | 0.1226 | – | – | – |
| Humans | – | 0.1165 | – | – | – |
| Species | 0 | 0.9929 | 94.90 | 1.8861 | 0 |
| Aerotolerance level | 0.46 | 0.4996 | 89.89 | 0.4847 | 0 |
Notes: *No result for chicken as it is treated as a reference group.
τ2: tau-squared; I2: inconsistency index; Q: Cochran’s Q statistic; R2: between-study variability.
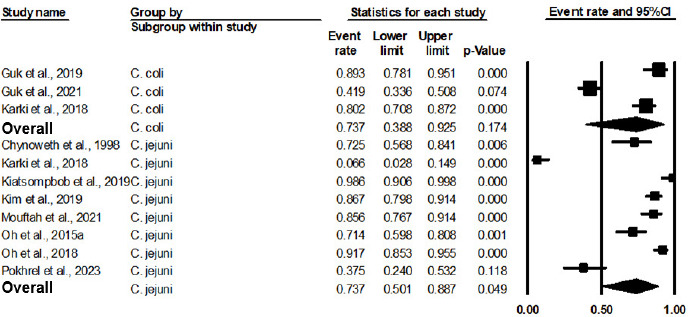
As shown in Figure 4 and Table 3, the pooled prevalence of AT C. jejuni was 73.7% (95% CI: 50.1% to 88.7%, P = 0.049). In comparison, the pooled prevalence of AT C. coli was statistically insignificant (P = 0.174), with an estimate of 73.7% (95% CI: 38.8% to 92.5%). Meta-regression in Table 4 reveals no significant difference between AT C. jejuni and AT C. coli, with P = 0.9929. However, C. jejuni was reported to have a higher survival bacterial concentration than C. coli after aerobic incubation (Mouftah et al., 2021).
Figure 4 Forest plot of the estimated prevalence of aerotolerant Campylobacter spp. by subgroup of the species.
As shown in Figure 5 and Table 3, the pooled prevalence of HAT Campylobacter spp. was 32.9% (95% CI: 22.6% to 45.1%, P = 0.007), with a Q statistic value of 100.20 and I2 of 91.02%. Similarly, the pooled prevalence of AT Campylobacter spp. was statistically significant (P = 0.028), with a pooled prevalence of 38.4% (95% CI: 29.0% to 48.7%). Meta-regression in Table 4 shows no significant difference between HAT and AT, with P = 0.4996. However, some studies reported a higher prevalence of HAT than AT isolated from animals, such as broilers and cattle, suggesting that HAT strains have an advantage of colonizing in the gastrointestinal tracts (GIT) of animals (Guk et al., 2019; Jaakkonen et al., 2020; Mouftah et al., 2021).
Figure 5 Forest plot of the estimated prevalence of aerotolerant Campylobacter spp. by the subgroup of aerotolerance level.
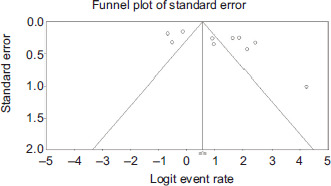
An I2 value of more than 75% indicated considerable heterogeneity (Higgins et al., 2003). Most of the heterogeneities in subgroup analysis and residual heterogeneities of all moderators in meta-regression were high (I2 > 88%), as shown in Table 4, which indicates the data synthesis should be interpreted with caution. These could be due to unmeasured covariates, such as the sampling and diagnostic techniques, season, temperature, and other underlying factors (Imrey et al., 2020). This highlights the need for future studies with a sample size (>30) that adopt a standardized aerotolerance assay. Further, there was a weak relationship between the prevalence and the source of isolates (between-study variability (R2) = 35.0%, P = 0.1497). In addition, no relationships were discovered regarding the publication year (R2 = 0% and P = 0.5653), location (R2 = 0% and P = 0.7839), species (R2 = 0% and P = 0.9929), and aerotolerance level (R2 = 0% and P = 0.4996) on the pooled Campylobacter spp. prevalence. Funnel test asymmetry (Figure S2) was confirmed by Egger’s test (P = 0.043), as shown in Table 5, revealing significant publication bias in this study. However, the Fail-safe N test found that the number of studies needed to revert the significance was 252. This value was much larger than the number of studies of 60, which was obtained from the formula of 5 k + 10. Thus, the result of the meta-analysis can be considered robust to publication bias (Rosenthal, 1979).
Table 5 Publication bias of meta-analysis.
| Number of studies | Egger’s test | 5 k +10 | Fail-safe N | ||
|---|---|---|---|---|---|
| Intercept | P value | ||||
| Overall | 10 | 7.26 | 0.043 | 60 | 232 |
The available prevalence data for AT Campylobacter spp., at the recent time point, are limited to C. jejuni and C. coli. Studies on the prevalence of other AT Campylobacter spp., such as C. concisus, C. upsaliensis, C. fetus, C. ureolyticus, and C. hyointestinalis, are the research gaps worth investigating, as these species are clinically important (Platts-Mills and Kosek, 2014). The prevalence of AT Campylobacter spp. in other retail foods is valuable, as Campylobacter spp. have been found in milk, eggs, fresh products, ready-to-eat food, and processed meat (Chai et al., 2007; Jaakkonen et al., 2020; Parisi et al., 2015; Toyomaki et al., 2012; Trimoulinard et al., 2017).
Survivability of aerotolerant Campylobacter spp. against environmental stress
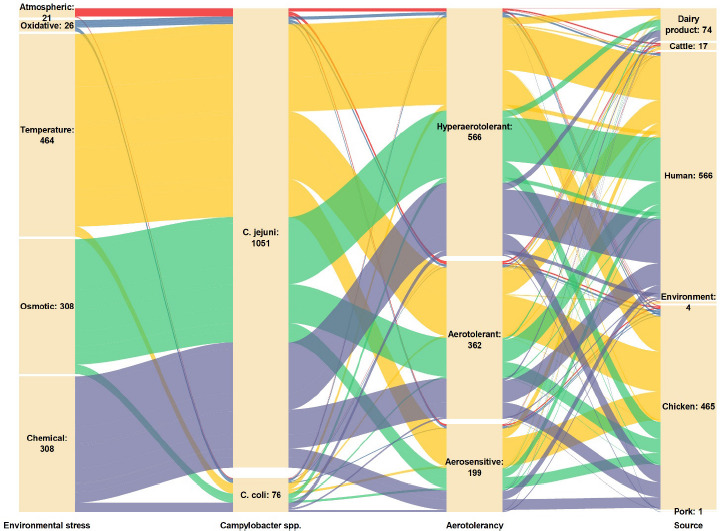
The survivability of AT Campylobacter spp. against environmental stresses is one of the forefront traits for the pathogen to induce campylobacteriosis in humans. Figure 6 displays the alluvial plot of the overview of the Campylobacter spp. isolates with different levels of aerotolerance studied against environmental stresses, as summarized in 17 studies (Chynoweth et al., 1998; Hur et al., 2024; Jaakkonen et al., 2020; Jones et al., 1993; Karki et al., 2018, 2019; Lee et al., 2019; Mouftah et al., 2021; O’Kane and Connerton, 2017; Oh et al., 2015a, 2017, 2018, 2019; Ortega-Sanz et al., 2024; Pokhrel et al., 2023; Rodrigues et al., 2015; Shagieva et al., 2021).
Figure 6 Alluvial plot of the overview of the isolates of Campylobacter spp. with different levels of aerotolerance from different sources studied against environmental stresses. The taller nodes indicate a larger number of isolates studied, with the number indicating the number of isolates.
The alluvial plot in Figure 6 demonstrates that AT C. jejuni (1,051 strains) was the most widely studied isolate against environmental stresses. Each of the environmental stresses are discussed further in the following subtopics. The alluvial plot displays that HAT Campylobacter spp. (566/1,127) were the most widely distributed strains compared to AT (362/1,127) and OS (199/1,127). Thus, it was postulated that aerotolerant (HAT and AT) Campylobacter spp. are becoming more ubiquitous than the OS strains.
Survivability of aerotolerant Campylobacter spp. against atmospheric and oxidative stresses
One important question that remains to be solved is how this ubiquitous yet microaerophilic and fastidious pathogen can thrive in aerobic environments and subsequently infect humans. Despite exposure to the aerobic atmosphere being closely associated with oxidative stress, the stress induced by aerobic incubation was referred to as aerobic stress in this study. In contrast, the stress caused by oxidative inducers was considered oxidative stress. Oxidative inducers included, such as hydrogen peroxide (H2O2), cumene hydroperoxide (CHP), menadione (MND), paraquat (PQ), are stated in Table S3. Tables S3 and S4 summarize the survival of Campylobacter spp. with different levels of aerotolerance under atmospheric and oxidative stress.
Hyperaerotolerant C. jejuni survived better than the AT strains under aerobic stress (Jaakkonen et al., 2020; Oh et al., 2017). Further, HAT C. jejuni were found to survive better than the AT strains, whereas AT survived better than the OS strains under different atmospheric conditions (CO2 and N2) in poultry meat. Thus, both HAT and AT strains might be able to thrive on the modified atmospheric packaging, further compromising food safety.
The most common Campylobacter strain used for studying atmospheric survivability was C. jejuni NCTC 11168 (Atack et al., 2008; Jaakkonen et al., 2020; Karki et al., 2019; O’Kane and Connerton, 2017; Oh et al., 2015b; Rodrigues et al., 2015). Jaakkonen et al. (2020) and Karki et al. (2019) reported contradicting results, as C. jejuni NCTC 11168 was reported as HAT and OS, respectively, based on the aerotolerance assay proposed by Oh et al. (2015a). However, several researchers investigated the aerotolerance of C. jejuni NCTC 11168 without concluding the AT classification (Atack et al., 2008; Gundogdu et al., 2015; Oh et al., 2015b; Rodrigues et al., 2015). Thus, the inconsistency of result and the lack of clarity of aerotolerance classification reaffirm the importance of standardized protocol.
Karki et al. (2019) found that retail beef meat juice, chicken liver juice, and beef liver juice contain factors that enhance the aerobic survival of C. jejuni and C. coli (Karki et al., 2019). This is because the heme-containing proteins in meat and liver juice act as cofactors for catalase and superoxide dismutase (Pretorius et al., 2016; van Vliet et al., 2002). In addition, iron in meat and liver juice mediates genes (fdxA and ahpC) and regulatory proteins (PerR and Fur) that are involved in aerotolerance mechanisms (Baillon et al., 1999; Butcher et al., 2015; van Vliet et al., 1999, 2001). Thus, C. jejuni and C. coli might survive better in situ than in vitro, and the actual prevalence of AT Campylobacter spp. could be higher before being detected in the laboratory.
The aerobic survival of C. jejuni cocktail (mixture of four strains) was investigated on different food matrices (chicken nuggets and minced meat) (Chynoweth et al., 1998). In sterile minced chicken, incubation at 25°C showed better survival than at 37°C. The higher survival at 25°C could be due to the pathogen being less metabolically active than at 37°C and thus less susceptible to reactive oxidative species (ROS) (Chynoweth et al., 1998). Further, the survival of C. jejuni in chicken nuggets was lower than in minced meat, possibly because of the seasonings and food additives in the nuggets that inhibit the growth of C. jejuni (Chynoweth et al., 1998). Thus, the survival of single and multiple strains of Campylobacter spp. in different food matrices is worth elucidating.
Hwang et al. (2014) was the only study to report the survival of non-jejuni/coli Campylobacter isolates under aerobic incubation. The researchers reported that C. fetus was able to survive in brain heart infusion after 12 h of aerobic incubation. However, the researchers did not state the incubation temperature and the classification for aerotolerance. This study demonstrated that Campylobacter spp. other than C. jejuni and C. coli might have aerotolerance properties.
Campylobacter spp. is normally exposed to ROS during colonization on a host, thriving in the environment, and within its cellular metabolism (Gundogdu et al., 2015). Several researchers investigated the effect of ROS-promoting agents (oxidative inducers) on the viability of AT Campylobacter spp., as shown in Table S4. Oh et al. (2015a) reported that aerotolerant C. jejuni (HAT and AT strains) survived significantly better than the non-AT C. jejuni (OS strains) against 1-mM H2O2, 100-µM CHP, and 100-µM MND, respectively. However, results from Ortega-Sanz et al. (2024) did not show any trend in aerotolerance and survival against oxidative stress. In comparison between C. jejuni and C. coli, Karki et al. (2018) reported that there were significant differences in H2O2 sensitivity between strains, while there was no correlation between H2O2 sensitivity and the level of aerotolerance (five C. jejuni and eight C. coli strains).
As shown in Table S4, Rodrigues et al. (2015) conducted an oxidative stress assay under MAC, while Karki et al. (2018) and Oh et al. (2015a) did not mention incubation atmosphere. It is essential to note that aerobic incubation might have acted synergistically with oxidative inducers on the survival of Campylobacter spp., potentially interfering with the observations.
Results from several research groups demonstrated that AT Campylobacter spp. had better survival and growth under atmospheric and oxidative stress. However, based on the paucity of studies investigating the relationship between aerotolerance, atmospheric stress, and oxidative stress defenses, the interaction between phenotypes remains inconclusive.
Aerobically acclimatized Campylobacter spp.: Aerobic acclimatization refers to the incubation of Campylobacter spp. under aerobic conditions fo r a defined period, allowing them to adapt to aerobic stress. Researchers conducted aerobic acclimatization differently, with incubation temperatures of 37°C or 42°C under aerobic conditions for 1–7 days and ranging from 1 to 15 aerobic passages on various agars (Chynoweth et al., 1998; Jones et al., 1993; O’Kane and Connerton, 2017; Rodrigues et al., 2015; Shagieva et al., 2021). It was observed that aerobically acclimatized C. jejuni survived longer on blood agar than non-aerobically acclimatized C. jejuni at 4°C under aerobic incubation (Jones et al., 1993). Similarly, O’Kane and Connerton (2017) reported that aerobic acclimatization improved the aerotolerance of C. coli OR12. Further, Shagieva et al. (2021) reported that aerobically acclimatized C. jejuni demonstrated aerobic growth on agar. Contrarily, aerobic acclimatization did not influence the growth and survivability of both C. jejuni Bf (Bf is a C. jejuni strain named by Rodrigues et al., 2015) and NCTC 11351 under aerobic conditions (Chynoweth et al., 1998; Rodrigues et al., 2015). These conflicting results underscore the importance of a standardized method for aerobic acclimatization.
It has been postulated that the aerotolerance of C. jejuni could be due to transcriptional and translational changes (Bronnec et al., 2016a, 2016b). Microaerobic passage did not affect C. coli OR12’s ability to grow in aerobic conditions, suggesting that aerotolerance is stable and is due to genetic modification in lieu of temporary physiological adaptation (O’Kane and Connerton, 2017). However, Nennig et al. (2022) reported that more than half (43/80) of C. jejuni were able to be aerobically acclimatized, and whole genome sequencing data suggested that genetic sequence does not govern this trait. Thus, the molecular mechanism of aerobic acclimatization and aerotolerance in Campylobacter spp. could be an interesting research question.
Growth of aerotolerant Campylobacter spp. under aerobic conditions: To our knowledge, Chynoweth et al. (1998) first reported a qualitative investigation on C. jejuni to grow on solid agar, and Rodrigues et al. (2015) were the first to report this quantitatively. C. jejuni Bf exhibited a stationary phase of 10 h, which could be regarded as the adaptive period for aerobic stress, followed by a growth of 2.5 log10 Colony Forming Units per milliliter (CFU/mL) from 10 h to 48 h on Kamali agar under aerobic incubation (Rodrigues et al., 2015). To date, no Campylobacter spp. other than C. jejuni and C. coli, have been reporte d to have the ability to grow under aerobic conditions, as shown in Table S5.
Aerobic growth of AT Campylobacter spp. is facilitated in liquid media (k H = 9.28 x 10-4 mol L-1 atm-1 in water at 42°C) because of lower oxygen transfer, compared to solid agar (Rodrigues et al., 2015). Next, a cell density of more than 104 CFU/mL could grant better aerobic stress tolerance in AT Campylobacter spp. because the oxygen demand was higher than that of low cell densities (lower than 104 CFU/mL) (Rodrigues et al., 2015). These two factors reduced the available oxygen and facilitated the aerobic growth of AT Campylobacter spp. (Rodrigues et al., 2015).
Shagieva et al. (2021), O’Kane and Connerton (2017), and Rodrigues et al. (2015) reported that Campylobacter spp. grew on Kamali, blood, and charcoal cefoperazone deoxycholate agars (CCD), respectively. These agar plates contain substances that neutralize toxic substances from oxygen (Gharst et al., 2013). However, the aerobic growth of C. jejuni on nutrient agar, as demonstrated by Chynoweth et al. (1998), might have greater aerobic growth potential because the agar did not contain any additives that mitigate aerobic stress.
Although the AT C. coli OR12 Aer P32 strain demonstrated aerobic growth on blood agar, it did not exhibit superior survival in liquid medium under aerobic conditions, unlike other C. jejuni and C. coli strains (O’Kane and Connerton, 2017). This finding did not justify such coherence, and the researchers did not explain it in their studies. Thus, this finding marks the importance of comparing the aerotolerance levels of Campylobacter spp. both in vitro (in broth and on agar) and in situ (food matrices).
Survivability of aerotolerant Campylobacter spp. against chemical antimicrobial agent
The only reported chemical antimicrobial agent used to control Campylobacter spp. in relation to aerotolerance is peracetic or peroxyacetic acid (PAA). PAA is the most commonly used antimicrobial in pre- and post-chill stages in poultry processing (Kataria et al., 2020). PAA offers a wide range of antimicrobial activities because of the combined action of acidic and oxidizing properties (Bauermeister et al., 2008). PAA is normally applied at main poultry chiller for up to 220 ppm, and for the post-chill immersion tank, the maximum permissible concentration is 2,000 ppm (United States Department of Agriculture-Food Safety and Inspection Service, 2019). PAA is an effective antimicrobial agent that reduces the population of Campylobacter spp. with a reduction of up to 1.3 log10 CFU (Chen et al., 2014). Only three studies were available on raw chicken skin, as summarized in Table S6. These studies found the same trend, with HAT strains survived better than the AT strains, whereas both HAT and AT strains thrived better than OS strains (Mouftah et al., 2021; Oh et al., 2018, 2019).
Most of C. jejuni isolates resisted PAA better than C. coli, with a higher final mean cell number (Mouftah et al., 2021). However, this is the only study that compared the survival of C. jejuni and C. coli. Since the application of PAA is in the equilibrium of H2O2 and acetic acid, the survival of HAT and AT strains in the presence of PAA could be due to the augmented oxidative stress defense (Oh et al., 2019; Yuan et al., 1997). Studies on the survivability of AT Campylobacter spp. against other chemical decontamination agents are the knowledge gaps that remain to be explored. Examples of other chemical decontamination agents used in Campylobacter spp. decontamination include chlorine, chlorine dioxide, acidified sodium chlorite, trisodium phosphate, and peroxy acid (Hansson et al., 2018).
Survivability of aerotolerant Campylobacter spp. against temperature stress
Temperature treatments, such as refrigeration, freezing, and pasteurization, are vital for food preservation. The survival of AT Campylobacter spp. against temperature stress is shown in Table S7. In situ studies reported that HAT strains survived better under refrigeration (4°C) than AT strains, whereas AT strains survived better than OS strains (Jaakkonen et al., 2020; Mouftah et al., 2021; Oh et al., 2017, 2018). These studies reported a consistent trend between aerotolerance levels and refrigeration survival. In addition, OS strains have a significantly higher proportion in the cold-sensitive group compared to the cold-tolerant group (Hur et al., 2024). On the contrary, Pokhrel et al. (2023) reported that AT strains survived better than HAT and OS strains in refrigerated chicken drumsticks. In comparison, in vitro studies produced mixed results. Lee et al. (2019) and Shagieva et al. (2021) found no difference between aerotolerance and refrigeration survival in MH broth and brain heart infusion broth, respectively. In contrast, C. jejuni OS strains reduced significantly more AT and HAT strains in both MH broth and chicken meat (Oh et al., 2017, 2019). Interestingly, Jaakkonen et al. (2020) reported variability in the refrigerated raw milk survival of C. jejuni within CC-21, even within the same ST, implying unconserved traits in survivability.
Similar to refrigeration, HAT Campylobacter spp. isolates portrayed significantly better survival on chicken skin than AT strains under freeze-thaw stress, whereas AT strains outperformed OS strains (Mouftah et al., 2021; Oh et al., 2018). Further, freeze-thaw stress in MH broth also showed a cognate trend as on chicken skin (Oh et al., 2019). On the contrary, Pokhrel et al. (2023) reported that AT strains survived better than HAT and OS strains in frozen chicken drumsticks. The discrepancy between studies could be due to the low number of HAT strains (five strains) in the study conducted by Pokhrel et al. (2023).
Under pasteurization (72°C, 15 s) and the extended heat-treatment (72°C, 30 s) in milk, HAT C. jejuni and C. coli strains survived better than AT strains, while OS survived the least (Mouftah et al., 2021; Oh et al., 2018). On the contrary, C. jejuni isolated from retail chicken were sensitive to heat (72°C, 30 s), regardless of AT properties (Oh et al., 2019). Varied survival trends of Campylobacter spp. with different aerotolerance levels against refrigeration, freeze-thaw, and heat treatment affirm the need for future study.
Similar to the aerobic stress trend described in the previous section, meat and liver juice significantly improved the survival of AT C. coli and non-AT C. jejuni at 4°C, compared to MH broth (Karki et al., 2019). It is conjectured that retail meat and liver juice from both chicken and beef had a significant influence, but not the origin of the juice. As a result, there could be better low-temperature survival of Campylobacter spp. under in situ conditions than under laboratory conditions.
Genes encoding cold shock proteins are not present in Campylobacter spp. (Hazeleger et al., 1998). Thus, Campylobacter spp. may harbor another tolerance mechanism in response to cold shock (Mouftah et al., 2021). Since some studies found a relation between aerotolerance and temperature stress survival, the mechanism behind this could be the existence and expression of certain genes that contribute to both phenotypes.
Survivability of aerotolerant Campylobacter spp. against osmotic stress
Campylobacter spp. are sensitive to hyperosmotic conditions, with inhibition reported at more than 2% NaCl, a common preservative used in food against pathogens (Doyle and Glass, 2010; Park, 2002). Only three studies have linked the aerotolerance of Campylobacter spp. to osmotic stress resistance, and all the studies were conducted in MH agar or MH broth, as shown in Table S8. Mouftah et al. (2021) and Oh et al. (2018) reported higher survival in HAT and AT C. jejuni or C. coli strains than OS strains under hyperosmotic stress (4% NaCl). Cross-protection might exist between aerotolerance and osmotic stress resistance, and vice versa (Mouftah et al., 2021). Conversely, the degree of hyper-osmotolerance was found to be greatly variable between strains and did not pertain to aerotolerance properties (Oh et al., 2019).
Half of the C. jejuni survived at 4% NaCl, whereas none of the C. coli survived, and difference in the final cell concentration was significant between the two species (Mouftah et al., 2021). However, only one study compared the two species. The relationship between aerotolerance and hyper-osmotolerance remains to be clarified through additional research, such as investigations of different food matrices and other Campylobacter spp.
Survivability of aerotolerant Campylobacter spp. against acidic stress
To the best of our knowledge, no study has elucidated the relationship between aerotolerance of Campylobacter spp. and acidic stress tolerance. Compared to other foodborne pathogens, C. jejuni is more sensitive to acid (Birk et al., 2010). This sensitivity is due to the lack of an acid resistance system (Birk et al., 2012; Merrell and Camilli, 1999; Park et al., 1996; Richard and Foster, 2003). However, Campylobacter spp. proteins, such as SodB, AhpC, and Dps, observed during acid stress, were associated with oxidative stress (Baillon et al., 1999; Birk et al., 2012; Ishikawa et al., 2003; Pesci et al., 1994; Purdy et al., 1999). Thus, C. jejuni might normally be in an oxygen-alert state, allowing it to remove ROS and undesirable components from acid stress (Afriliana et al., 2018; Birk et al., 2012).
The pH of certain fermented foods is in the range of 4.5–5.5 (Nout, 1994). In addition, the pH of coffee ranges from 5.4 to 6.2, while the pH of milk is 6.5–6.7 (Afriliana et al., 2018; Renan et al., 2006; Sikand et al., 2010). Another example of acidic food is marinated meat (Birk et al., 2010). AT Campylobacter spp. could survive better in acidic food, which poses a food safety issue. The prevalence and survivability of AT Campylobacter spp. in the aforementioned acidic food are worth investigating.
Influence of atmospheric conditions on the survivability of aerotolerant Campylobacter spp. against environmental stresses
Certain survival studies against oxidative, temperature, and osmotic stresses, as shown in Tables S4, S7, and S8, respectively, did not provide information on the atmospheric incubation conditions. Atmospheric conditions could influence the survival mechanism of Campylobacter spp. against environmental stresses. For instance, AT and HAT C. jejuni showed better resistance to different atmospheric stresses (aerobic, N2, and CO2) than AT, whereas AT strains had better resistance than OS strains at 4°C (Oh et al., 2017). Furthermore, C. jejuni (with all levels of aerotolerance) survived significantly better in refrigeration under aerobic conditions than microaerobic conditions (Hur et al., 2024). Thus, atmospheric conditions, such as aerobic and modified atmospheric packaging, on the survival of AT Campylobacter spp. against environmental stresses, is an interesting research gap.
Inactivation dynamics of aerotolerant Campylobacter spp.
Different experimental settings of the included studies in Section “Survivability of aerotolerant Campylobacter spp. against environmental stress” render data harmonization difficult. Some studies reported the results in duration and percentage of survival, while others presented the results in charts without numeric figures. Thus, based on the available numeric data, the inactivation dynamics are summarized as follows:
-
Under aerobic stress, HAT, AT, and OS C. jejuni had 2, 3, and 4 log10 CFU reduction, respectively, after 3 days of incubation in MH broth (Oh et al., 2017). Further, AT C. coli declined to below the limit of detection (1.3 log10 CFU/mL) from inoculation of 6 log10 CFU/mL after 24 h aerobic incubation in MH broth (O’Kane and Connerton, 2017).
-
Under oxidative stress, 3.35 µM CHP exposure for 60 min reduced viability of AT and HAT C. jejuni > 6.25 ± 0.32 log10 CFU/mL, while 5 mM H2O2 reduced 5.89 ± 0.60 log10 CFU/mL (Ortega-Sanz et al., 2024). Under 0.5 mM PQ exposure for 60 min, AT C. jejuni reduced 1.4–1.5 log10 CFU/mL (Rodrigues et al., 2015).
-
Under chemical antimicrobial stress, PAA reduced HAT and AT C. jejuni and C. coli equally by ca. 4.5 log10 CFU/mL, while OS strains were reduced by ca. 7 log10 CFU/mL (Mouftah et al., 2021). Furthermore, PAA inactivated HAT and AT C. jejuni by ca. 2.1 and 2.5 log10 CFU/mL, respectively, while OS strains were reduced by 3.7 log10 CFU/mL (Oh et al., 2019).
-
Under chilling (4°C) stress, storing HAT, AT, and OS C. jejuni for 7 days reduced 4.9, 5.1, and 7.4 log10 CFU/mL, respectively (Oh et al., 2019). On contrary, at the same condition as Oh et al. (2019), greater reduction was reported in HAT C. jejuni (0.27 log10 CFU/mL) than OS and AT C. jejuni with 0.14 log10 CFU/mL and 0.08 log10 CFU/mL, respectively. After 5 days of chilling, OS C. jejuni reduced 1.5 log10 CFU/g and AT C. jejuni reduced 2.1–3.0 log10 CFU/g (Lee et al., 2019).
-
Under freezing (-20°C) stress, HAT, OS, and AT C. jejuni reduced 1.12, 0.90, and 0.60 log10 CFU/mL, respectively, regardless of the storage period (Pokhrel et al., 2023).
Gene regulation of aerotolerant Campylobacter spp.
Gene regulation underlying AT Campylobacter spp. for environmental persistence is intricate and remains poorly understood. AT C. jejuni under aerobic incubation and after aerobic acclimatization was reported to have an increase in the transcription levels of genes involved in aerobic detoxification, which include sodB, ahpC, katA, tpx, and trxB (Rodrigues et al., 2016). In relation to the elevated transcription levels, the corresponding proteins (AhpC, KatA, Tpx, and TrxB) were reported to have higher abundance after aerobic incubation (Rodrigues et al., 2016). Similarly, katA, ahpC, and trxB were reported to have higher expression in aerobic incubation than microaerobic conditions in HAT C. coli, whereas their expression decreased in the OS C. coli (Guk et al., 2022). Further, ahpC mutation in HAT C. jejuni reduced aerotolerance level to AT (Oh et al., 2015a). AT C. jejuni was reported with higher expression in cosR (oxidative stress response), oorDABC (oxygen metabolism), and mreB (cell morphology) after aerobic incubation (Bronnec et al., 2016b; Rodrigues et al., 2015). Under aerobic incubations at 37°C and 4°C, the expression of htrB (stress response) gene was higher in AT C. jejuni than in OS C. jejuni (Lee et al., 2019). Further studies are warranted to elucidate the gene regulatory mechanisms of AT Campylobacter spp. in response to environmental stress.
Mechanism of persistence of aerotolerant Campylobacter spp.
C. jejuni devolves into viable but non-culturable (VNBC) stage and forms biofilms upon aerobic stress (Mouftah et al., 2021; Oh et al., 2015b; Yagi et al., 2022). Yagi et al. (2022) reported that the AT C. jejuni strain remained in VBNC stage for a longer period than the aero-sensitive strains under aerobic stress. The researchers also postulated that AT strains might enter the VBNC stage more slowly than the aero-sensitive strains under aerobic conditions (Yagi et al., 2022). This might be due to the AT strains resisting aerobic stress more, and the longer VBNC stage implies greater environmental persistence. In addition, Oh et al. (2015b) postulated that AT C. jejuni enters the VBNC stage after 12 h of aerobic incubation. However, the research did not compare to the aero-sensitive strain.
Aerotolerance was found to have a relationship with biofilm formation potential in C. jejuni and C. coli, with HAT isolates showing significantly greater biofilm formation potential than AT and OS isolates at 42°C under microaerobic conditions (Mouftah et al., 2021). On the contrary, Pokhrel et al. (2024) reported that different aerotolerance levels yielded no significant difference in the number of C. jejuni biofilms attached to stainless steel coupons at 42°C, regardless of the microaerobic and aerobic conditions. However, at room temperature, a significantly lower number of C. jejuni biofilm attached to stainless steel coupons was observed in HAT C. jejuni than AT and OS counterparts (Pokhrel et al., 2024). This suggests that incubation temperature could influence the formation of biofilms. In addition, Ortega-Sanz et al. (2024) reported that significantly higher biofilm formation was observed in HAT C. jejuni than in AT strains under aerobic conditions on polystyrene, but no significant difference was observed on stainless steel coupons. Thus, the type of material might influence the formation of biofilm. Further, AT C. jejuni Bf strain developed similar biofilm volume and thickness under both aerobic and microaerobic conditions (Bronnec et al., 2016b). However, under aerobic incubation, a more compact and structured biofilm was formed, creating a microaerobic niche that supported the growth (Bronnec et al., 2016b).
HAT and AT C. jejuni are found to have higher superoxide dismutase and catalase activities (the two important enzymes in oxidative stress defense) than OS strains (Oh et al., 2019). C. jejuni possesses both surface polysaccharides, capsular polysaccharide (CPS) and lipopolysaccharide (LPS), on the outer membrane (Jeon et al., 2009). Under aerobic conditions, the carbon metabolism of C. jejuni increased, leading to elevated amino acid uptake (Kim et al., 2023). This stimulated the formation of thicker CPS and LPS, which potentially function as a permeability barrier, protecting the pathogen from excess oxygen in aerobic atmosphere (Kim et al., 2023). Thus, the comparison of the structure of CPS and LPS among aerotolerance levels could be interesting. Further, the relationship between aerotolerance, VBNC, biofilm, oxidative stress defense enzymes, and protective polysaccharides of Campylobacter spp. remains scanty.
Genetic relatedness of aerotolerant Campylobacter spp. strains
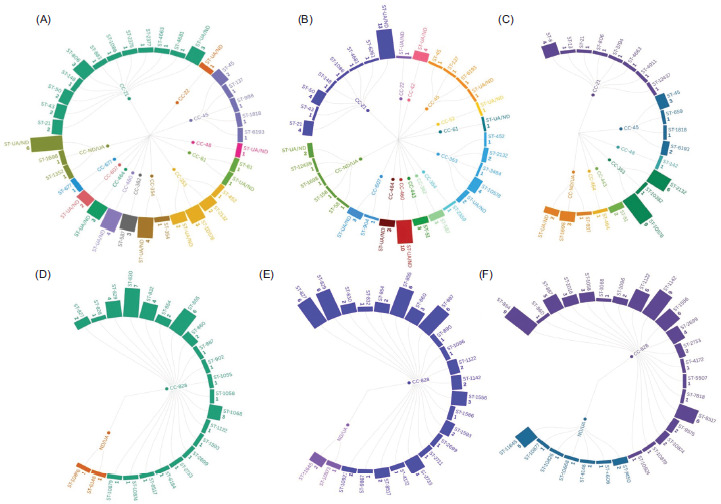
Discriminatory typing methods for Campylobacter spp. are important for improving the understanding of epidemiology and genetic background (Nielsen et al., 2010). Multilocus sequence typing (MLST) is a valuable typing tool for discriminating Campylobacter spp. isolates and defining population structure (Dingle et al., 2001, 2005). The ST obtained from MLST can be grouped into CC based on relatedness (Nielsen et al., 2010). Table S9 shows the summary of eight studies on the classification of CC and ST based on aerotolerance level in C. jejuni and C. coli. The aerotolerance level comparison between ST was not made for Kiatsomphob et al. (2019), as the study did not disclose ST for each CC. Data from Table S9 are regrouped based on the respective CC and ST of C. jejuni and C. coli with aerotolerance levels, and are shown in Figure 7. Data from certain studies that did not classify CC to aerotolerance were further analyzed and classified based on the data from their published articles and supplementary documents (Guk et al., 2019, 2021; Jaakkonen et al., 2020; Kiatsomphob et al., 2019).
Figure 7 Radial graphs of CC and ST of (A) OS C. jejuni, (B) AT C. jejuni, (C) HAT C. jejuni, (D) OS C. coli, (E) AT C. coli, and (F) HAT C. coli. The frequency of ST is in bold face below the respective ST. CC: clonal complex; ST: sequence type; UA: unassigned to any CC; ND: not determined; OS: aero-sensitive; AT: aerotolerant; HAT: hyperaerotolerant.
Ordinal logistic regression analysis of 364 Campylobacter spp. isolates (input data from Table S9) showed that ST had a statistically significant (P < 0.001) influence on aerotolerance level. It has been postulated that the ST of Campylobacter spp. is related to aerotolerance levels. As shown in Figure 7, CC-353 showed the highest prevalence in HAT C. jejuni, while CC-21 showed the highest prevalence in AT C. jejuni. CC-353 was associated with a very high level of resistance to erythromycin (Jehanne et al., 2025). Further, CC-21 was the most prevalent in human and poultry slaughterhouse (Jeong et al., 2025).
In C. coli, CC-828 was the most prevalent CC in all three classes of aerotolerance (OS, AT, and HAT) strains, as shown in Figure 7. The most prevalent C. coli ST among both HAT and AT in the study of Guk et al. (2019) was ST-855, and this ST was not detected in OS strains. In contrast, the highest prevalent ST (ST-1068) among HAT strains in the study conducted by Guk et al. (2021) was detected in OS strains. Further, MLST data of C. coli isolates from ducks were compared to genetic relatedness to human sources in PubMLST and NCBI databases (Guk et al., 2019). Eight ST out of 18 from duck isolates were identified and found to be shared with human sources (Guk et al., 2019). HAT C. coli had a significantly higher proportion among shared ST than non-shared ST, while OS showed the opposite trend (Guk et al., 2019). In addition, a higher proportion of genetic relatedness was also reported between swine-derived HAT C. coli and human C. coli isolates, than OS strains (Guk et al., 2021).
Several studies reported a high proportion of shared STs in Campylobacter spp. between food-producing animals and human clinical cases (Asakura et al., 2019; Litrup et al., 2007; Ramonaite et al., 2017). Findings of Ortega-Sanz et al. (2024), Kiatsomphob et al. (2019), and Rodrigues et al. (2015), as summarized in Table S9, showed that HAT and AT strains were highly prevalent in human campylobacteriosis cases. Thus, the relationship between aerotolerance level and the overlap ST between food-producing animals and human clinical cases will be an interesting research field.
Risk of aerotolerant Campylobacter spp. to humans
In view of the enhanced resistance of AT Campylobacter spp. against multiple environmental stresses, such as atmospheric stress, disinfectant exposure, freeze-thaw, refrigeration, heat treatment, and osmotic stress, the AT Campylobacter spp. strains might survive better throughout the food chain. This would, in turn, augment foodborne transmission to humans (Guk et al., 2019; Mouftah et al., 2021).
Mihaljevic et al. (2007) found that 5 h aerobic exposure of C. jejuni significantly elevated invasion capacity in the Caco-2 cells model, suggesting that normal atmospheric conditions might be associated with the pathogenicity of pathogen. The AT Campylobacter spp. might have better oxidative stress resistance and could become more pathogenic, as the pathogen might be less susceptible to the free radicals produced by host immune response (Atack and Kelly, 2009). Furthermore, the aerotolerance of Campylobacter spp. is to some extent related to oxidative stress resistance and colonization factors in chickens, which contribute to their high prevalence in food as well as their elevated pathogenicity in human infections (Bolton, 2015; Flint et al., 2016; Hermans et al., 2011).
The risk of AT Campylobacter spp. to human health can be higher than that of the normal Campylobacter spp. evaluated in the past, as AT strains are commonly present in food. Thus, stricter regulations could be implemented for AT Campylobacter spp., as in the case of E. coli O157:H7 in the pathogenic E. coli strain.
Antibiotic resistance in aerotolerant Campylobacter spp.
To date, studies relating the aerotolerance profile of Campylobacter spp. to the antibiotic resistance profile are scarce, with no study conducted on C. jejuni. No significant differences were found between the antibiotic resistance rates and aerotolerance profiles in 56 C. coli isolates (Guk et al., 2019). However, 25% (7/28) of HAT and 9.1% (2/22) of AT C. coli isolates were highly resistant to ciprofloxacin (≥32 µg/mL), while none was from OS isolates (Guk et al., 2019). Similarly, in another study conducted by the same research group on pig feces, HAT C. coli showed the highest proportion (18.8%) in high-level ciprofloxacin resistance (Guk et al., 2021). Thus, AT strains might be more difficult to treat and place a greater medical burden because of higher antibiotic resistance. However, more studies are warranted to elucidate the relationship between aerotolerance and antibiotic resistance profiles.
Prevalence of virulence genes in aerotolerant Campylobacter spp.
Similar to the antibiotic-resistant profile, only a few studies linked virulence genes to the aerotolerance of Campylobacter spp. HAT C. coli isolates were reported to possess all 10 virulence genes analyzed, such as flaA (motility), flhB (motility), cadF (adhesion), pldA (adhesion), iamA (invasion), ceuE (invasion), cdtA (cytoxin), wlaN (Guillain–Barré syndrome), hcp (type VI secretion system), and virB11 (type IV secretion system), with the prevalence ranging from 3.6% to 100% (Guk et al., 2019, 2021). As compared to HAT C. coli, only six virulence genes were reported in OS C. coli, whereas seven virulence genes were found in AT C. coli (Guk et al., 2019). However, the prevalence of virulence genes was not significantly different between OS, AT, and HAT isolates (Guk et al., 2019, 2021).
Conversely, among 70 C. jejuni isolates, HAT C. jejuni strains depicted significantly higher frequencies of virulence genes than OS strains, suggesting HAT strains could be more pathogenic to humans than their OS counterparts (Oh et al., 2017). Similarly, Rodrigues et al. (2018) reported that AT C. jejuni Bf strain is one of the most virulent strains among the 10 strains tested. However, the study did not relate the aerotolerance levels of strains to the level of virulence.
The gene involved in the aerobic survival of Campylobacter spp. can also contribute to virulence. For instance, sodB, which is important for C. coli survival under aerobic conditions, plays an important role in colonizing the chick (Purdy et al., 1999). In the context of the virulence genes expression, such as cadF, cdtB (cytotoxin), ciaB (invasion), and clpP (stress tolerance), no significant difference was discovered in the expression between two AT C. jejuni strains and one OS C. jejuni strain (Lee et al., 2019). Therefore, the prevalence and expression of virulence genes in relation to aerotolerance level warrants further study to improve our understanding. AT strains with higher virulence genes might increase pathogenicity and thus contribute to higher campylobacteriosis cases.
Conclusion and the Way Forward
Research has increasingly demonstrated that AT Campylobacter spp. are prevalent in retail food, exhibiting increased pathogenicity and resistance to multiple environmental stresses. AT Campylobacter spp. might bring immense burdens to the economy and health care. The persistence of AT Campylobacter spp. is associated with a battery of survival mechanisms against various environmental stresses. Based on the current findings, it will not be surprising that the term ‘aerobic microorganism’ could be used to describe Campylobacter spp. in near future. This demonstrated the dire need to bolster our understanding of the relationship between survivability and the associated mechanisms in order to develop a holistic preventive measure to curtail this emerging pathogen.
To translate the available findings into real-world food safety practices, a surveillance program for AT Campylobacter spp. in the food chain is pivotal, and policymakers should urgently evaluate and revise the current protocol for public health. From that, modified atmospheric packaging, better hygienic practices, and greater coverage of cold chain transportation could be corrective measures to be considered.
In order to better contribute to future research in AT Campylobacter spp., several research gaps could be underpinned as illustrated in Figure S3 and stated as follows:
-
Standardized protocols for aerotolerance level classification and aerobic acclimatization.
-
Prevalence study of aerotolerance level and aerobic growth involving a larger sample size, a variety of foods, and different geographical regions.
-
Elucidation on other clinically important Campylobacter spp.
-
Survivability of single-strain, multiple-strain of the same species and multiple species against environmental stresses in vitro and in situ.
-
Identification of ST of the strains and relating it to the source of the isolate and aerotolerance level.
-
Phenotypic mechanism for aerotolerance, aerobic acclimatization, and aerobic growth.
-
Antibiotic resistance profile on a variety of antibiotics with different doses and the prevalence and expression of virulence genes.
-
Gene regulatory mechanism between aerotolerance, antibiotic resistance, and virulence.
-
Characterization and assessment of health burden via risk assessment for better policy implementation.
Mandatory Disclosure on Use of Artificial Intelligence
The authors declare that no AI-assisted tools were used in the preparation of this manuscript. All references have been manually verified for accuracy and relevance.
Declaration of Completing Interest
The authors declared that no completing financial interests or personal relationship that could influence the work of this study involved.
Acknowledgement
The authors would like to thank the Ministry of Higher Education Malaysia for the Higher Institution Centre of Excellence (HICoE) grant scheme (grant No.: UPM.ITAFoS.800–3/8/HICoE (FasaII)/2023/5230004). The authors also thanked the laboratory assistants of the Faculty of Food Science and Technology for their kind assistance.
Author Contributions
All authors contributed equally to this article.
Conflict of Interest
The authors declared no conflict of interest.
REFERENCES
Afriliana, A., Harada, H., and Khotijah, P.Q. 2018. Fermented technology of robusta coffee beans (Canephora coffee) with kefir milk to produce specialty coffee. In: 4th International Conference on Food, Agriculture and Natural Resources. Atlantis Press, Dordrecht, the Netherlands, pp. 301–308. 10.2991/fanres-18.2018.61
Asakura, H., Sakata, J., Nakamura, H., Yamamoto, S., and Murakami, S. 2019. Phylogenetic diversity and antimicrobial resistance of Campylobacter coli from humans and animals in Japan. Microbes and Environments 34: 146–154. 10.1264/jsme2.ME18115
Atack, J.M., Harvey, P., Jones, M.A., and Kelly, D.J. 2008. The Campylobacter jejuni thiol peroxidases Tpx and Bcp both contribute to aerotolerance and peroxide-mediated stress resistance but have distinct substrate specificities. Journal of Bacteriology 190: 5279–5290. 10.1128/JB.00100-08
Atack, J.M., and Kelly, D.J. 2009. Oxidative stress in Campylobacter jejuni: responses, resistance and regulation. Future Microbiology 4: 677–690. 10.2217/fmb.09.44
Backert, S., Tegtmeyer, N., Cróinín, T.Ó., Boehm, M., and Heimesaat, M.M. 2017. Human campylobacteriosis. In: G. Klein (ed.) Campylobacter. Academic Press, Cambridge, MA, pp. 1–25.
Baek, Y.J., Song, J.E., Kim, E.J., Choi, H., Sohn, Y., Jeon, Y.D., Lee, E.H., Ahn, J.Y., Jeong, S.J., Ku, N.S., Choi, J.Y., Yeom, J.S., Song, Y.G., and Kim, J.H. 2024. Trends, clinical characteristics, antimicrobial susceptibility patterns, and outcomes of Campylobacter bacteraemia: a multicentre retrospective study. Infection 52(3): 857–864. 10.1007/s15010-023-02118-4
Baillon, M.-L.A., van Vliet, A.H., Ketley, J.M., Constantinidou, C., and Penn, C.W. 1999. An iron-regulated alkyl hydroperoxide reductase (AhpC) confers aerotolerance and oxidative stress resistance to the microaerophilic pathogen Campylobacter jejuni. Journal of Bacteriology 181: 4798–4804.
Bauermeister, L.J., Bowers, J.W., Townsend, J.C., and McKee, S.R. 2008. Validating the efficacy of peracetic acid mixture as an antimicrobial in poultry chillers. Journal of Food Protection 71: 1119–1122. 10.4315/0362-028X-71.6.1119
Begley, M., and Hill, C. 2015. Stress adaptation in foodborne pathogens. Annual Review of Food Science and Technology 6: 191–210. 10.1146/annurev-food-030713-092350
Birk, T., Grønlund, A.C., Christensen, B.B., Knøchel, S., Lohse, K., and Rosenquist, H. 2010. Effect of organic acids and marination ingredients on the survival of Campylobacter jejuni on meat. Journal of Food Protection 73: 258–265. 10.4315/0362-028X-73.2.258
Birk, T., Wik, M.T., Lametsch, R., and Knøchel, S. 2012. Acid stress response and protein induction in Campylobacter jejuni isolates with different acid tolerance. BMC Microbiology 12: 1–13. 10.1186/1471-2180-12-174
Bolton, D.J. 2015. Campylobacter virulence and survival factors. Food Microbiology 48: 99–108. 10.1016/j.fm.2014.11.017
Borenstein, M., Hedges, L.V., Higgins, J.P., and Rothstein, H.R. 2009. Introduction to Meta-Analysis. John Wiley, West Sussex, UK.
Borenstein, M. 2022. Comprehensive Meta-Analysis Software. In: M. Egger, J.P.T. Higgins and G. Davey Smith (eds.) Systematic Reviews in Health Research. John Wiley & Sons Ltd, Oxford, UK, pp. 109-128. 10.1002/9781119099369.ch27
Bronnec, V., Haddad, N., Cruveiller, S., Hernould, M., Tresse, O., and Zagorec, M. 2016a. Draft genome sequence of Campylobacter jejuni Bf, an atypical strain able to grow under aerobiosis. Genome Announcements 4: e00120–00116. 10.1128/genomeA.00120-16
Bronnec, V., Turoňová, H., Bouju, A., Cruveiller, S., Rodrigues, R., Demnerova, K., Tresse, O., Haddad, N., and Zagorec, M. 2016b. Adhesion, biofilm formation, and genomic features of Campylobacter jejuni Bf, an atypical strain able to grow under aerobic conditions. Frontiers in Microbiology 7: 1002. 10.3389/fmicb.2016.01002
Bücker, R., Krug, S., Moos, V., Bojarski, C., Schweiger, M., Kerick, M., Fromm, A., Janssen, S., Fromm, M., and Hering, N. 2018. Campylobacter jejuni impairs sodium transport and epithelial barrier function via cytokine release in human colon. Mucosal Immunology 11: 474–485. 10.1038/mi.2017.66
Butcher, J., Handley, R.A., van Vliet, A.H., and Stintzi, A. 2015. Refined analysis of the Campylobacter jejuni iron-dependent/independent Fur-and PerR-transcriptomes. BMC Genomics 16: 1–13. 10.1186/s12864-015-1661-7
Centers for Disease Control and Prevention (CDC) 2024. Campylobacter infection (Campylobacteriosis). Available at: www.cdc.gov/campylobacter/index.html (Accessed: 10 July 2024).
Chai, L.C., Robin, T., Ragavan, U.M., Gunsalam, J.W., Bakar, F.A., Ghazali, F.M., Radu, S., and Kumar, M.P. 2007. Thermophilic Campylobacter spp. in salad vegetables in Malaysia. International Journal of Food Microbiology 117: 106–111. 10.1016/j.ijfoodmicro.2007.02.014
Chen, X., Bauermeister, L.J., Hill, G.N., Singh, M., Bilgili, S.F., and McKee, S.R. 2014. Efficacy of various antimicrobials on reduction of Salmonella and Campylobacter and quality attributes of ground chicken obtained from poultry parts treated in a postchill decontamination tank. Journal of Food Protection 77: 1882–1888. 10.4315/0362-028X.JFP-14-114
Churchill, K.J., Sargeant, J.M., Farber, J.M., and O’Connor, A.M. 2019. Prevalence of Listeria monocytogenes in select ready-to-eat foods—deli meat, soft cheese, and packaged salad: a systematic review and meta-analysis. Journal of Food Protection 82: 344–357. 10.4315/0362-028X.JFP-18-158
Chynoweth, R., Hudson, J., and Thom, K. 1998. Aerobic growth and survival of Campylobacter jejuni in food and stream water. Letters in Applied Microbiology 27: 341–344. 10.1046/j.1472-765X.1998.00453.x
Dingle, K.E., Colles, F.M., Falush, D., and Maiden, M.C. 2005. Sequence typing and comparison of population biology of Campylobacter coli and Campylobacter jejuni. Journal of Clinical Microbiology 43: 340–347. 10.1128/jcm.43.1.340-347.2005
Dingle, K., Colles, F., Wareing, D., Ure, R., Fox, A., Bolton, F., Bootsma, H., Willems, R., Urwin, R., and Maiden, M. 2001. Multilocus sequence typing system for Campylobacter jejuni. Journal of Clinical Microbiology 39: 14–23. 10.1128/jcm.39.1.14-23.2001
Doyle, M.E., and Glass, K.A. 2010. Sodium reduction and its effect on food safety, food quality, and human health. Comprehensive Review of Food Science and Food Safety 9: 44–56. 10.1111/j.1541-4337.2009.00096.x
Egger, M., Smith, G.D., Schneider, M., and Minder, C. 1997. Bias in meta-analysis detected by a simple, graphical test. British Medical Journal 315: 629–634. 10.1136/bmj.315.7109.629
European Food Safety Authority (EFSA) and European Centre for Disease Prevention and Control (ECDC). 2024. The European Union One Health 2023 zoonoses report. EFSA Journal 22: e9106. 10.2903/j.efsa.2024.9106
Flint, A., Butcher, J., and Stintzi, A. 2016. Stress responses, adaptation, and virulence of bacterial pathogens during host gastrointestinal colonization. Microbiology Spectrum 4: 385–411. 10.1128/microbiolspec.VMBF-0007-2015
Gharst, G., Oyarzabal, O.A., and Hussain, S.K. 2013. Review of current methodologies to isolate and identify Campylobacter spp. from foods. Journal of Microbiological Methods 95: 84–92. 10.1016/j.mimet.2013.07.014
Guk, J.-H., Kim, J., Song, H., Kim, J., An, J.-U., Kim, J., Ryu, S., Jeon, B., and Cho, S. 2019. Hyper-aerotolerant Campylobacter coli from duck sources and its potential threat to public health: virulence, antimicrobial resistance, and genetic relatedness. Microorganisms 7: 579. 10.3390/microorganisms7110579
Guk, J.-H., Song, H., Yi, S., An, J.-U., Lee, S., Kim, W.-H., and Cho, S. 2021. Hyper-aerotolerant Campylobacter coli from swine may pose a potential threat to public health based on its quinolone resistance, virulence potential, and genetic relatedness. Frontiers in Microbiology 12: 703993. 10.3389/fmicb.2021.703993
Guk, J.H., Woo, J., Song, H., Kim, W.H., Kim, J., Ryu, S., and Cho, S. 2022. Hyper-aerotolerant Campylobacter coli, an emerging foodborne pathogen, shows differential expressions of oxidative stress-related genes. Veternary Microbiology 264: 109308. 10.1016/j.vetmic.2021.109308
Gundogdu, O., da Silva, D.T., Mohammad, B., Elmi, A., Mills, D.C., Wren, B.W., and Dorrell, N. 2015. The Campylobacter jejuni MarR-like transcriptional regulators RrpA and RrpB both influence bacterial responses to oxidative and aerobic stresses. Frontiers in Microbiology 6: 724. 10.3389/fmicb.2015.00724
Haines, M., Eberle, K., McDaniel, C., and Kiess, A. 2011. Evaluating 3 gas-delivery systems for culturing Campylobacter jejuni in a microaerophilic environment. Poultry Science 90: 2378–2382. 10.3382/ps.2011-01463
Hanna, J., Neill, S.D., O’brien, J.J., and Ellis, W.A. 1983. Comparison of aerotolerant and reference strains of Campylobacter species by polyacrylamide gel electrophoresis. International Journal of Systematic and Evolutionary Microbiology 33: 143–146. 10.1099/00207713-33-2-143
Hansson, I., Sandberg, M., Habib, I., Lowman, R., and Engvall, E.O. 2018. Knowledge gaps in control of Campylobacter for prevention of campylobacteriosis. Transboundary and Emerging Diseases 65: 30–48. 10.1111/tbed.12870
Hazeleger, W.C., Wouters, J.A., Rombouts, F.M., and Abee, T. 1998. Physiological activity of Campylobacter jejuni far below the minimal growth temperature. Applied Environmental Microbiology 64: 3917–3922. 10.1128/AEM.64.10.3917-3922.1998
Heimesaat, M.M., Backert, S., Alter, T., and Bereswill, S. 2023. Molecular targets in Campylobacter infections. Biomolecules 13(3): 409. 10.3390/biom13030409
Hermans, D., Van Deun, K., Martel, A., Van Immerseel, F., Messens, W., Heyndrickx, M., Haesebrouck, F., and Pasmans, F. 2011. Colonization factors of Campylobacter jejuni in the chicken gut. Veterinary Research 42: 1–14. 10.1186/1297-9716-42-82
Higgins, J.P., Thompson, S.G., Deeks, J.J., and Altman, D.G. 2003. Measuring inconsistency in meta-analyses. British Medical Journal 327(7414): 557–560. 10.1136/bmj.327.7414.557
Hur, J.I., Kim, J., Kang, M.S., Kim, H.J., Ryu, S., and Jeon, B. 2024. Cold tolerance in Campylobacter jejuni and its impact on food safety. Food Research International 175: 113683. 10.1016/j.foodres.2023.113683
Hwang, S., Miller, W.G., Ryu, S., and Jeon, B. 2014. Divergent distribution of the sensor kinase CosS in non-thermotolerant Campylobacter species and its functional incompatibility with the response regulator CosR of Campylobacter jejuni. PloS One 9: e89774. 10.1371/journal.pone.0089774
Imbrea, A.M., Balta, I., Dumitrescu, G., McCleery, D., Pet, I., Iancu, T., Stef, L., Corcionivoschi, N., and Liliana, P.C. 2024. Exploring the contribution of Campylobacter jejuni to post-infectious irritable bowel syndrome: a literature review. Applied Sciences 14(8): 3373. 10.3390/app14083373
Imrey, P.B. 2020. Limitations of meta-analyses of studies with high heterogeneity. JAMA Network Open 3(1): Article e1919325. 10.1001/jamanetworkopen.2019.19325
Ishikawa, T., Mizunoe, Y., Kawabata, S.-i., Takade, A., Harada, M., Wai, S.N., and Yoshida, S.-i. 2003. The iron-binding protein Dps confers hydrogen peroxide stress resistance to Campylobacter jejuni. Journal of Bacteriology 185: 1010–1017. 10.1128/jb.185.3.1010-1017.2003
Iversen, A., Rortveit, G., Wensaas, K.A., and Gulla, C.O. 2024. The impact on primary care of a large waterborne Campylobacter outbreak in Norway: a controlled observational study. Scandavian Journal of Primary Health Care 42(1): 187–194. 10.1080/02813432.2023.2299116
Jaakkonen, A., Kivistö, R., Aarnio, M., Kalekivi, J., and Hakkinen, M. 2020. Persistent contamination of raw milk by Campylobacter jejuni ST-883. PloS One 15: e0231810. 10.1371/journal.pone.0231810
Je, H.J., Kim, U.I., and Koo, O.K. 2024. A comprehensive systematic review and meta-analysis of Listeria monocytogenes prevalence in food products in South Korea. International Journal of Food Microbiology 415: 110655. 10.1016/j.ijfoodmicro.2024.110655
Jehanne, Q., Bénéjat, L., Ducournau, A., Aptel, J., Pivard, M., Gillet, L., Jauvain, M., and Lehours, P. 2025. Increasing rates of erm (B) and erm (N) in human Campylobacter coli and Campylobacter jejuni erythromycin-resistant isolates between 2018 and 2023 in France. Antimicrobial Agents and Chemotherapy 69(2): e01668–24. 10.1128/aac.01668-24
Jeon, B., Muraoka, W., Scupham, A., and Zhang, Q. 2009. Roles of lipooligosaccharide and capsular polysaccharide in antimicrobial resistance and natural transformation of Campylobacter jejuni. Journal of Antimicrobial Chemotherapy 63: 462–468. 10.1093/jac/dkn529
Jeong, J., Lee, J.Y., Moon, J.S., Kang, M.S., Kang, S.I., Lee, O.M., Lee, S.H, Kwon, Y.K., Chae, M., and Cho, S. 2025. Virulence genes, antimicrobial resistance, and genotypes of Campylobacter jejuni isolated from chicken slaughterhouses in South Korea. Foodborne Pathogens and Disease 22(4): 281–289. 10.1089/fpd.2023.0144
Jones, D., Sutcliffe, E., Rios, R., Fox, A., and Curry, A. 1993. Campylobacter jejuni adapts to aerobic metabolism in the environment. Journal of Medical Microbiology 38: 145–150. 10.1099/00222615-38-2-145
Kaakoush, N.O., Miller, W.G., De Reuse, H., and Mendz, G.L. 2007. Oxygen requirement and tolerance of Campylobacter jejuni. Research in Microbiology 158: 644–650. 10.1016/j.resmic.2007.07.009
Kanters, S. 2022. Fixed- and random-effects models. In: Evangelou, E., and Veroniki, A.A. (eds.) Meta-Research. Methods in Molecular Biology. Humana, New York, NY, vol 2345, pp. 41–65. 10.1007/978-1-0716-1566-9_3
Karki, A.B., Marasini, D., Oakey, C.K., Mar, K., and Fakhr, M.K. 2018. Campylobacter coli from retail liver and meat products is more aerotolerant than Campylobacter jejuni. Frontiers in Microbiology 9: 2951. 10.3389/fmicb.2018.02951
Karki, A.B., Wells, H., and Fakhr, M.K. 2019. Retail liver juices enhance the survivability of Campylobacter jejuni and Campylobacter coli at low temperatures. Scientific Reports 9: 2733. 10.1038/s41598-018-35820-7
Kataria, J., Vaddu, S., Rama, E.N., Sidhu, G., Thippareddi, H., and Singh, M. 2020. Evaluating the efficacy of peracetic acid on Salmonella and Campylobacter on chicken wings at various pH levels. Poultry Science 99: 5137–5142. 10.1016/j.psj.2020.06.070
Keithlin, J., Sargeant, J., Thomas, M.K., and Fazil, A. 2014. Systematic review and meta-analysis of the proportion of Campylobacter cases that develop chronic sequelae. BMC Public Health 14: 1–19. 10.1186/1471-2458-14-1203
Kelly, P., and Hodges, P. 2024. Infectious diarrhoea. Medicine 52(4): 197–203. 10.1016/j.mpmed.2024.01.008
Kiatsomphob, S., Taniguchi, T., Tarigan, E., Latt, K.M., Jeon, B., and Misawa, N. 2019. Aerotolerance and multilocus sequence typing among Campylobacter jejuni strains isolated from humans, broiler chickens, and cattle in Miyazaki prefecture, Japan. Journal of Veternary Medical Sciences 81: 1144–1151. 10.1292/jvms.19-0228
Kim, J., Park, M., Ahn, E., Mao, Q., Chen, C., Ryu, S., and Jeon, B. 2023. Stimulation of surface polysaccharide production under aerobic conditions confers aerotolerance in Campylobacter jejuni. Microbiology Spectrum 11: e03761–03722. 10.1128/spectrum.03761-22
Kim, J., Park, H., Kim, J., Kim, J.H., Jung, J.I., Cho, S., Ryu, S., and Jeon, B. 2019. Comparative analysis of aerotolerance, antibiotic resistance, and virulence gene prevalence in Campylobacter jejuni isolates from retail raw chicken and duck meat in South Korea. Microorganisms 7: 433. 10.3390/microorganisms7100433
Lee, H., Lee, S., Kim, S., Ha, J., Lee, J., Choi, Y., Oh, H., Kim, Y., Lee, Y., and Yoon, Y. 2019. The risk of aerotolerant Campylobacter jejuni strains in poultry meat distribution and storage. Microbial Pathogenesis 134: 103537. 10.1016/j.micpath.2019.05.020
Litrup, E., Torpdahl, M., and Nielsen, E. 2007. Multilocus sequence typing performed on Campylobacter coli isolates from humans, broilers, pigs and cattle originating in Denmark. Journal of Applied Microbiology 103: 210–218. 10.1111/j.1365-2672.2006.03214.x
Lynch, Ó.A., Cagney, C., McDowell, D.A., and Duffy, G. 2011. Occurrence of fastidious Campylobacter spp. in fresh meat and poultry using an adapted cultural protocol. International Journal of Food Microbiology 150: 171–177. 10.1016/j.ijfoodmicro.2011.07.037
Merrell, D.S., and Camilli, A. 1999. The cadA gene of Vibrio cholerae is induced during infection and plays a role in acid tolerance. Molecular Microbiology 34: 836–849. 10.1046/j.1365-2958.1999.01650.x
Mihaljevic, R.R., Sikic, M., Klancnik, A., Brumini, G., Mozina, S.S., and Abram, M. 2007. Environmental stress factors affecting survival and virulence of Campylobacter jejuni. Microbial Pathogenesis 43: 120–125. 10.1016/j.micpath.2007.03.004
Ministry of Health Singapore 2025. Communicable Diseases Surveillance in Singapore 2021–2022. Available at: https://doi.org/isomer-user-content.by.gov.sg/3/afc27f07-74af-44a9-a8e1-0050249e8860/Chapter%204_%20Foodborne%20diseases.pdf (Accessed: 5 May 2025).
Mouftah, S.F., Cobo-Díaz, J.F., Álvarez-Ordóñez, A., Mousa, A., Calland, J.K., Pascoe, B., Sheppard, S.K., and Elhadidy, M. 2021. Stress resistance associated with multi-host transmission and enhanced biofilm formation at 42°C among hyper-aerotolerant generalist Campylobacter jejuni. Food Microbiology 95: 103706. 10.1016/j.fm.2020.103706
Myintzaw, P., Jaiswal, A.K., and Jaiswal, S. 2023. A review on campylobacteriosis associated with poultry meat consumption. Food Reviews International 39(4): 2107–2121. 10.1080/87559129.2021.1942487
Neill, S., Ellis, W., and O’brien, J. 1979. Designation of aerotolerant Campylobacter-like organisms from porcine and bovine abortions to the genus Campylobacter. Research in Veterinary Science 27: 180–186. 10.1016/s0034-5288(18)32825-x
Nennig, M., Clément, A., Longueval, E., Bernardi, T., Ragimbeau, C., and Tresse, O. 2022. Metaphenotypes associated with recurrent genomic lineages of Campylobacter jejuni responsible for human infections in Luxembourg. Frontiers in Microbiology 13: 901192. 10.3389/fmicb.2022.901192
Nielsen, L.N., Sheppard, S., McCarthy, N., Maiden, M., Ingmer, H., and Krogfelt, K. 2010. MLST clustering of Campylobacter jejuni isolates from patients with gastroenteritis, reactive arthritis and Guillain–Barré syndrome. Journal of Applied Microbiology 108: 591–599. 10.1111/j.1365-2672.2009.04444.x
Nout, M. 1994. Fermented foods and food safety. Food Research International 27: 291–298. 10.1016/0963-9969(94)90097-3
Oh, E., Andrews, K.J., McMullen, L.M., and Jeon, B. 2019. Tolerance to stress conditions associated with food safety in Campylobacter jejuni strains isolated from retail raw chicken. Scientific Reports 9: 11915. 10.1038/s41598-019-48373-0
Oh, E., Chui, L., Bae, J., Li, V., Ma, A., Mutschall, S.K., Taboada, E.N., McMullen, L.M., and Jeon, B. 2018. Frequent implication of multistress-tolerant Campylobacter jejuni in human infections. Emerging Infectious Diseases 24: 1037. 10.3201/eid2406.171587
Oh, E., McMullen, L.M., Chui, L., and Jeon, B. 2017. Differential survival of hyper-aerotolerant Campylobacter jejuni under different gas conditions. Frontiers in Microbiology 8: 954. 10.3389/fmicb.2017.00954
Oh, E., McMullen, L., and Jeon, B. 2015a. High prevalence of hyper-aerotolerant Campylobacter jejuni in retail poultry with potential implication in human infection. Frontiers in Microbiology 6: 1263. 10.3389/fmicb.2015.01263
Oh, E., McMullen, L., and Jeon, B. 2015b. Impact of oxidative stress defense on bacterial survival and morphological change in Campylobacter jejuni under aerobic conditions. Frontiers in Microbiology 6: 295. 10.3389/fmicb.2015.00295
O’Kane, P.M., and Connerton, I.F. 2017. Characterisation of aerotolerant forms of a robust chicken colonizing Campylobacter coli. Frontiers in Microbiology 8: 513. 10.3389/fmicb.2017.00513
Ortega-Sanz, I., Bocigas, C., Melero, B., and Rovira, J. 2024. Phase variation modulates the multi-phenotypes displayed by clinical Campylobacter jejuni strains. Food Microbiology 117: 104397. 10.1016/j.fm.2023.104397
Ouzzani, M., Hammady, H., Fedorowicz, Z., & Elmagarmid, A. (2016). Rayyan—a web and mobile app for systematic reviews. Systematic reviews 5(1): 210. 10.1186/s13643-016-0384-4
Page, M.J., McKenzie, J.E., Bossuyt, P.M., Boutron, I., Hoffmann, T.C., Mulrow, C.D, Shamseer, L., Tetzlaff, J.M., Akl, E.A., Brennan, S.E., and Chou, R., 2021. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. British Medical Journal 372: n71. 10.1136/bmj.n71
Parisi, M., Northcutt, J., Smith, D., Steinberg, E., and Dawson, P. 2015. Microbiological contamination of shell eggs produced in conventional and free-range housing systems. Food Control 47: 161–165. 10.1016/j.foodcont.2014.06.038
Park, S.F. 2002. The physiology of Campylobacter species and its relevance to their role as foodborne pathogens. International Journal of Food Microbiology 74: 177–188. 10.1016/S0168-1605(01)00678-X
Park, Y.K., Bearson, B., Bang, S.H., Bang, I.S., and Foster, J.W. 1996. Internal pH crisis, lysine decarboxylase and the acid tolerance response of Salmonella typhimurium. Molecular Microbiology 20: 605–611. 10.1046/j.1365-2958.1996.5441070.x
Pesci, E.C., Cottle, D.L., and Pickett, C.L. 1994. Genetic, enzymatic, and pathogenic studies of the iron superoxide dismutase of Campylobacter jejuni. Infection and Immunity 62: 2687–2694. 10.1128/iai.62.7.2687-2694.1994
Platts-Mills, J.A., and Kosek, M. 2014. Update on the burden of Campylobacter in developing countries. Current Opinion in Infectious Diseases 27: 444–450. 10.1097/QCO.0000000000000091
Plishka, M., Sargeant, J.M., Greer, A.L., Hookey, S., and Winder, C. 2021. The prevalence of Campylobacter in live cattle, Turkey, chicken, and swine in the United States and Canada: a systematic review and meta-analysis. Foodborne Pathogens and Disease 18: 230–242. 10.1089/fpd.2020.2834
Pokhrel, D., Thames, H.T., Fugate, H., Dinh, T., Schilling, W., White, S., Ramachandran, R., Sukumaran, A.T., and Zhang, L. 2024. Increase in temperature facilitates Campylobacter jejuni biofilm formation under both aerobic and microaerobic incubation. Poultry Science 103: 103753. 10.1016/j.psj.2024.103753
Pokhrel, D., Thames, H.T., Zhang, L., Dinh, T., Schilling, M.W., White, S., Ramachandran, R., and Sukumaran, A.T. 2023. Aerotolerance and multi-locus sequence typing of Campylobacter jejuni isolated from commercial broiler processing plants. Foods 12: 3305. 10.3390/foods12173305
Pretorius, B., Schönfeldt, H.C., and Hall, N. 2016. Total and haem iron content lean meat cuts and the contribution to the diet. Food Chemistry 193: 97–101. 10.1016/j.foodchem.2015.02.109
Purdy, D., Cawthraw, S., Dickinson, J.H., Newell, D.G., and Park, S.F. 1999. Generation of a superoxide dismutase (SOD)-deficient mutant of Campylobacter coli: evidence for the significance of SOD in Campylobacter survival and colonization. Applied Environmental Microbiology 65: 2540–2546. 10.1128/AEM.65.6.2540-2546.1999
Ramonaite, S., Tamuleviciene, E., Alter, T., Kasnauskyte, N., and Malakauskas, M. 2017. MLST genotypes of Campylobacter jejuni isolated from broiler products, dairy cattle and human campylobacteriosis cases in Lithuania. BMC infectious Diseases 17(1): 430. 10.1186/s12879-017-2535-1
Renan, M., Mekmene, O., Famelart, M.-H., Guyomarc’h, F., Arnoult-Delest, V., Pâquet, D., and Brulé, G. 2006. pH-dependent behaviour of soluble protein aggregates formed during heat-treatment of milk at pH 6.5 or 7.2. Journal of Dairy Research 73: 79–86. 10.1017/S0022029905001627
Richard, H.T., and Foster, J.W. 2003. Acid resistance in Escherichia coli. Advances in Applied Microbiology 52: 167–186. 10.1016/s0065-2164(03)01007-4
Rodrigues, R.C., Haddad, N., Chevret, D., Cappelier, J.M., and Tresse, O. 2016. Comparison of proteomics profiles of Campylobacter jejuni strain Bf under microaerobic and aerobic conditions. Frontiers in Microbiology 7: 1596. 10.3389/fmicb.2016.01596
Rodrigues, R.C., Pocheron, A.-L., Cappelier, J.-M., Tresse, O., and Haddad, N. 2018. An adapted in vitro assay to assess Campylobacter jejuni interaction with intestinal epithelial cells: taking into stimulation with TNFα. Journal of Microbiological Methods 149: 67–72. 10.1016/j.mimet.2018.04.020
Rodrigues, R.C., Pocheron, A.-L., Hernould, M., Haddad, N., Tresse, O., and Cappelier, J.-M. 2015. Description of Campylobacter jejuni Bf, an atypical aero-tolerant strain. Gut Pathogens 7: 1–12. 10.1186/s13099-015-0077-x
Rosenthal, R. 1979. The file drawer problem and tolerance for null results. Psychological Bulletin 86: 638. 10.1037/0033-2909.86.3.638
Rothstein, H.R., Sutton, A.J., and Borenstein, M. 2005. Publication Bias in Meta-Analysis. John Wiley, Chichester, WSX, UK.
Shagieva, E., Demnerova, K., and Michova, H. 2021. Waterborne isolates of Campylobacter jejuni are able to develop aerotolerance, survive exposure to low temperature, and interact with acanthamoeba polyphaga. Frontiers in Microbiology 12: 730858. 10.3389/fmicb.2021.730858
Sheth, V.H., Shah, N.P., Jain, R., Bhanushali, N., and Bhatnagar, V. 2024. Development and validation of a risk-of-bias tool for assessing in vitro studies conducted in dentistry: the QUIN. Journal of Prosthetic Dentistry 131(6): 1038–1042. 10.1016/j.prosdent.2022.05.019
Sikand, V., Tong, P., and Walker, J. 2010. Heat stability of reconstituted, protein-standardized skim milk powders. Journal of Dairy Science 93: 5561–5571. 10.3168/jds.2010-3128
Song, H., Kim, J., Guk, J.-H., An, J.-U., Lee, S., and Cho, S. 2020. Complete genome sequence and comparative genomic analysis of hyper-aerotolerant Campylobacter lari strain SCHS02 isolated from duck for its potential pathogenicity. Microbial Pathogenesis. 142: 104110. 10.1016/j.micpath.2020.104110
Toyomaki, H., Mahundi, E., Ishihara, K., Kurwijila, L.R., Grace, D., and Makita, K. 2012. Quantitative risk assessment of acquiring campylobacteriosis from consumption of ready-to-eat beef in Arusha Municipality, Tanzania. Journal of Veternary Epidemiology 16: 31–32. 10.2743/jve.16.31
Trimoulinard, A., Beral, M., Henry, I., Atiana, L., Porphyre, V., Tessier, C., Leclercq, A., and Cardinale, E. 2017. Contamination by Salmonella spp., Campylobacter spp., and Listeria spp. of most popular chicken-and pork-sausages sold in Reunion Island. International Journal of Food Microbiology 250: 68–74. 10.1016/j.ijfoodmicro.2017.03.017
United States Department of Agriculture-Food Safety and Inspection Service 2019. Safe and suitable ingredients used in the production of meat, poultry, and egg products. Direc. 7120.1. Rev. 52. Available at: www.fsis.usda.gov/wps/wcm/connect/bab10e09-aefa-483b-8be8-809a1f051d4c/7120.1.pdf?MOD5AJPERES (Accessed: 10 October 2023).
Vandamme, P., Falsen, E., Rossau, R., Hoste, B., Segers, P., Tytgat, R., and De Ley, J. 1991. Revision of Campylobacter, Helicobacter, and Wolinella taxonomy: emendation of generic descriptions and proposal of Arcobacter gen. nov. International Journal of Systematic and Evolutionary Microbiology 41: 88–103. 10.1099/00207713-41-1-88
Vandamme, P., Vancanneyt, M., Pot, B., Mels, L., Hoste, B., Dewettinck, D., Vlaes, L., Van Den Borre, C., Higgins, R., and Hommez, J. 1992. Polyphasic taxonomic study of the emended genus Arcobacter with Arcobacter butzleri comb. nov., and Arcobacter skirrowii sp. nov., an aerotolerant bacterium isolated from veterinary specimens. International Journal of Systematic and Evolutionary Microbiology 42: 344–356. 10.1099/00207713-41-1-88
van Enst, W.A., Ochodo, E., Scholten, R.J., Hooft, L., and Leeflang, M.M. 2014. Investigation of publication bias in meta-analyses of diagnostic test accuracy: a meta-epidemiological study. BMC Medical Research Methodology 14: 1–11. 10.1186/1471-2288-14-70
van Vliet, A.H., Baillon, M.-L.A., Penn, C.W., and Ketley, J.M. 1999. Campylobacter jejuni contains two fur homologs: characterization of iron-responsive regulation of peroxide stress defense genes by the PerR repressor. Journal of Bacteriology 181: 6371–6376. 10.1128/JB.181.20.6371-6376.1999
van Vliet, A.H., Baillon, M.-L.A., Penn, C.W., and Ketley, J.M. 2001. The iron-induced ferredoxin FdxA of Campylobacter jejuni is involved in aerotolerance. FEMS Microbiology Letters 196: 189–193. 10.1111/j.1574-6968.2001.tb10563.x
van Vliet, A.H., Ketley, J.M., Park, S.F., and Penn, C.W. 2002. The role of iron in Campylobacter gene regulation, metabolism and oxidative stress defense. FEMS Microbiology Review 26: 173–186. 10.1111/j.1574-6976.2002.tb00609.x
Yagi, S., Okada, A., and Inoshima, Y. 2022. Role of temperature, nutrition, oxygen, osmolality, and bacterial strain in inducing a viable but non-culturable state in Campylobacter jejuni. Journal of Microbiological Methods 195: 106456. 10.1016/j.mimet.2022.106456
Yuan, Z., Ni, Y., and Van Heiningen, A. 1997. Kinetics of peracetic acid decomposition: part I: spontaneous decomposition at typical pulp bleaching conditions. Canadian Journal of Chemical Engineering 75: 37–41. 10.1002/cjce.5450750108
Supplementary
Table S1 Summary of risk of bias assessment (quality assessment) of 39 included articles.
| Clearly stated aims/ objectives | Detailed explanation of sampling technique | Details of comparison group | Detailed explanation of methodology | Randomization | Method of measurement of outcome | Statistical analysis | Presentation of results | Scores (%) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Atack et al., 2008 | 1 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 81.25 |
| 2 | Baillon et al., 1999 | 2 | 2 | 2 | 2 | 0 | 2 | 1 | 2 | 81.25 |
| 3 | Bronnec et al., 2016a | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 4 | Bronnec et al., 2016b | 2 | 2 | 2 | 2 | 0 | 2 | 1 | 2 | 81.25 |
| 5 | Chynoweth et al., 1998 | 2 | 2 | 2 | 2 | 0 | 2 | 1 | 2 | 81.25 |
| 6 | Guk et al., 2019 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 7 | Guk et al., 2021 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 8 | Guk et al., 2022 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 9 | Gundogdu et al., 2015 | 1 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 81.25 |
| 10 | Hur et al., 2024 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 11 | Hwang et al., 2014 | 1 | 2 | 2 | 2 | 0 | 2 | 1 | 2 | 75 |
| 12 | Jaakkonen et al., 2020 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 13 | Jones et al., 1993 | 2 | 0 | 2 | 2 | 0 | 2 | 1 | 2 | 68.75 |
| 14 | Kaakoush e t al., 2007 | 2 | 2 | 2 | 2 | 0 | 2 | 1 | 2 | 81.25 |
| 15 | Karki et al., 2018 | 2 | 2 | 2 | 2 | 0 | 2 | 1 | 2 | 81.25 |
| 16 | Karki et al., 2019 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 17 | Kiatsomphob et al., 2019 | 1 | 2 | 2 | 2 | 0 | 2 | 1 | 2 | 75 |
| 18 | Kim et al., 2019 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 19 | Kim et al., 2023 | 1 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 81.25 |
| 20 | Lee et al., 2019 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 21 | Mouftah et al., 2021 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 22 | Oh et al., 2015a | 1 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 81.25 |
| 23 | Oh et al., 2015b | 1 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 81.25 |
| 24 | Oh et al., 2017 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 25 | Oh et al., 2018 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 26 | Oh et al., 2019 | 1 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 81.25 |
| 27 | O’Kane and Connerton, 2017 | 1 | 2 | 2 | 2 | 0 | 2 | 1 | 2 | 75 |
| 28 | Ortega et al., 2024 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 29 | Pokhrel et al., 2023 | 1 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 81.25 |
| 30 | Pokhrel et al., 2024 | 2 | 2 | 2 | 2 | 0 | 2 | 1 | 2 | 81.25 |
| 31 | Purdy et al., 1999 | 1 | 2 | 2 | 2 | 0 | 2 | 1 | 2 | 75 |
| 32 | Rodrigues et al., 2015 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 33 | Rodrigues et al., 2016 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 34 | Rodrigues et al., 2018 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
| 35 | Shagieva et al., 2021 | 1 | 2 | 2 | 2 | 0 | 2 | 1 | 2 | 75 |
| 36 | Song et al., 2020 | 2 | 2 | 2 | 2 | 0 | 2 | 1 | 2 | 81.25 |
| 37 | van Vliet et al., 2001 | 1 | 2 | 2 | 2 | 0 | 2 | 1 | 2 | 75 |
| 38 | Vandamme et al., 1992 | 1 | 2 | 2 | 2 | 0 | 2 | 1 | 2 | 75 |
| 39 | Yagi et al., 2022 | 2 | 2 | 2 | 2 | 0 | 2 | 2 | 2 | 87.5 |
Figure S1 Schematic diagram of standardization protocol for aerotolerance assay.
Figure S2 Funnel plot assessing publication bias for prevalence of aerotolerant Campylobacter spp..
Table S2 Summary of sixteen articles on the prevalence of aerotolerant Campylobacter spp.
| Isolate origin | Aerotolerance of C. jejuni Percentage (number of isolates/ total number of isolates) | Aerotolerance of C. coli Percentage (number of isolates/ total number of isolates) | References | ||||
|---|---|---|---|---|---|---|---|
| OS | AT | HAT | OS | AT | HAT | ||
| Clinical sample | 0% (0/5) | 40% (2/5) | 60% (3/5) | – | – | – | Ortega–Sanz et al., 2024 |
| Poultry processing plant | 62.5% (25/40) | 25% (10/40) | 12.5% (5/40) | – | – | – | Pokhrel et al., 2023 |
| Swine faeces | – | – | – | 58.1% (72/124) | 28.2% (35/124) | 13.7% (17/124) | Guk et al., 2021 |
| Dairy product, broiler carcass, and clinical sample | 14.4% (13/90) | 20% (18/90) | 65.6% (59/90) | 23.8% (5/21) | 19.1% (4/21) | 57.1% (12/21) | Mouftah et al., 2021 |
| Pond water, outlet of wastewater treatment plant, raw chicken meat and clinical setting | 78.6% (11/14) | 21.4% (3/14) | –* | – | – | – | Shagieva et al., 2021 |
| Milk, cattle, and udder cloth | 0% (0/8) | 37.5% (3/8) | 62.5% (5/8) | – | – | – | Jaakkonen et al., 2020 |
| Duck carcass and meat | – | – | – | 10.7% (6/56) | 39.3% (22/56) | 50% (28/56) | Guk et al., 2019 |
| Chicken liver and human | 100% (2/2) | – | – | – | – | – | Karki et al., 2019 |
| Chicken liver and beef liver | – | – | – | 0%(0/2) | 50% (1/2) | 50% (1/2) | Karki et al., 2019 |
| Chicken meat | 13.3% (12/90) | 50% (45/90) | 36.7% (33/90) | – | – | – | Kim et al., 2019 |
| Duck meat | 13.3% (6/45) | 62.2% (28/45) | 24.4% (11/45) | – | – | – | Kim et al., 2019 |
| Human | 0% (0/26) | 84.6% (22/26) | 15.4% (4/26) | – | – | – | Kiatsomphob et al., 2019 |
| Broiler | 5% (1/20) | 35% (7/20) | 60% (12/20) | Kiatsomphob et al., 2019 | |||
| Cattle | 0% (0/24) | 50% (12/24) | 50% (12/24) | Kiatsomphob et al., 2019 | |||
| Chicken | 100% (8/8) | 0% (0/8) | –* | – | – | – | Lee et al., 2019 |
| Duck | 66.7% (4/6) | 33.3% (2/6) | |||||
| Human stool | 8.3% (10/121) | 38.0% (46/121) | 53.7% (65/121) | – | – | – | Oh et al., 2018 |
| Chicken meat, chicken liver, chicken gizzard, beef liver, turkey and pork | 93.4% (71/76) | 3.9% (3/76) | 2.6% (2/76) | 38.5% (18/91) | 19.8% (35/91) | 41.8% (38/91) | Karki et al., 2018 |
| Chicken meats | 28.6% (20/70) | 35.7% (25/70) | 35.7% (25/70) | – | – | – | Oh et al., 2015a |
| Human, water and raw chicken | 27.5% (11/40) | 72.5% (29/40) | –* | – | – | – | Chynoweth et al., 1998 |
| N/A | 0% (0/25) | 100% (25/25) | –* | Jones et al., 1993 | |||
OS = Aero-sensitive; AT = Aerotolerant; HAT = Hyper-aerotolerant; N/A = Not available; * = HAT classification not determined.
Table S3 Summary of six articles on the survival of Campylobacter spp. with different levels of aerotolerance under atmospheric stress.
| Aerotolerance level of isolates | Isolate origin | Number of isolates | Experimental Media | Incubating conditions | Findings | References |
|---|---|---|---|---|---|---|
| C. jejuni (HAT, AT) | Milk, cattle, human, udder cloth, and cattle | 6 HAT and 3 AT | MH broth | 106 CFU/mL with 200 rpm shaking at 41.5°C for 72 h under AC | • HAT C. jejuni survived longer than AT C. jejuni. | Jaakkonen et al., 2020 |
| C. coli (HAT, AT) and C. jejuni (OS) | Chicken liver, beef liver, and human | 1 HAT, 1 AT, and OS | 10% liver juice, 10% meat juice, MH broth, and 10% laked horse blood | OD600=0.2 in PBS with 200 rpm shaking at 42°C under AC | • Addition of beef liver juice, chicken liver juice, beef meat juice, and laked horse blood enhanced the aerotolerance. | Karki et al., 2019 |
| C. jejuni (ND), C. coli (ND, AT & aerobic passaged) | Human and chicken | 4 ND and 1 AT | MH broth | 6 log10 CFU/mL with 100 rpm shaking at 37°C for 30 h under AC | • Aerobic passaged C. coli did not survive better than its wild type. | O’Kane and Connerton, 2017 |
| C. jejuni (HAT, AT, and OS) | Poultry | 2 HAT, 2 AT, and 2 OS | Raw chicken meat and MH broth | OD600=0.1at 4°C under aerobic, CO2, and N2 conditions for 14 days | • Under aerobic incubation in meat, HAT and AT survived significantly higher than OS. HAT survived for 14 days while AT survived for 7 days. • Under CO2, and N2 incubation in meat, HAT and AT survived significantly better than OS in both CO2 and N2, respectively. HAT and AT survived for 14, days under N2, while OS survived for 7 days. HAT, AT, and OS survived for 7, 5, and 3 days under CO2 , respectively. • Under aerobic incubation in MH broth, HAT, AT, and OS had 2, 3, and 4 log10 CFU reduction, respectively within 3 days. HAT, AT, and OS strains survived for 7 days. |
Oh et al., 2017 |
| C. jejuni cocktails (AT) | Human, veterinary, and raw chicken | 4 AT | Raw mince chicken, autoclaved mince chicken, and chicken nugget | 106 CFU/g without shaking at 25°C or 37°C for 8 days under AC | • AT cocktails survived better at 25°C than at 37°C. | Chynoweth et al., 1998 |
| C. jejuni (AT) | 13997 (human) | 1 AT | BA | 9-10 log10 per plate without shaking at 42°C and 37°C for 34 days under AC and MAC | • AT strain survived longer under AC than MAC under both 37°C and 42°C. | Jones et al., 1993 |
OS = Aero-sensitive; AT = Aerotolerant; HAT = Hyper-aerotolerant; ND = Aerotolerant status not determined; AC = Aerobic condition; MAC = Microaerobic condition; MH = Mueller Hinton; BA = Blood agar.
Table S4 Summary four articles on the survival of Campylobacter spp. with different levels of aerotolerance under oxidative stress.
| Aerotolerance level of isolates | Isolate origin | Number of isolates | Experimental Media | Incubating conditions | Findings | References |
|---|---|---|---|---|---|---|
| C. jejuni (AT, HAT) | Clinical setting | 3 HAT and 2 AT | MH broth | 108 CFU/mL and exposed to 5 mM H2O2, 3.35 µM CHP at 37°C for 1 h (atmosphere not specified) | • One AT strain survived significantly less than the HAT strains, while the other AT strain survived significantly more than the HAT strains. | Ortega-Sanz et al., 2024 |
| C. coli (HAT, AT, OS) and C. jejuni (HAT, AT, OS) | Chicken meat, chicken liver, chicken gizzard, pork, and beef liver | 6 HAT, 4 AT, and 4 OS | MH agar | OD600 = 0.15 in PBS and exposed to 0.05, 0.1, and 0.5 mM H2O2 at 45°C for 24 h (atmosphere not specified) | • No correlation between H2O2 and aerotolerance. | Karki et al., 2018 |
| C. jejuni (HAT, AT, OS) | Retail broiler | 2 HAT, 2 AT, and 2 OS | MH broth | 108 CFU/mL and exposed to 1 mM H2O2, 100 µM CHP or 100 µM MND for 1 h (atmosphere and temperature not specified) | • OS strains survived significantly less than AT and HAT strains against H2O2, CHP, and MND. • AT strains survived significantly less than HAT strains against H2O2, CHP, and MND. |
Oh et al., 2015a |
| C. jejuni (ND, AT) | NCTC 11168 and human | 1 ND and 1 AT | Peptone saline solution | OD600 = 0.03 and exposed to 0.12, 0.25, 0.50, and 1 mM of H2O2 or PQ at 42°C for 1 h under MAC | • AT strain demonstrated a better survival rate than ND strain under H2O2 and PQ. | Rodrigues et al., 2015 |
OS = Aero-sensitive; AT = Aerotolerant; HAT = Hyper-aerotolerant; ND = Aerotolerant status not determined; H2O2 = Hydrogen peroxide; CHP = Cumene hydroperoxide; MND = Menadione; PQ = Paraquat; BA = Blood agar; MH = Mueller Hinton; AC = Aerobic condition; MAC = Microaerobic condition; PBS = Phosphate Buffered Saline.
Table S5 Summary of four articles on the aerobic growth of Campylobacter spp.
| Aerotolerance level of isolates | Isolate origin | Number of isolates | Experimental Media | Incubating conditions | Findings | References |
|---|---|---|---|---|---|---|
| C. jejuni (NA) | Pond water, outlet of wastewater treatment plant, raw chicken meat, and clinical setting | 14 | Kamali agar | 3.0 McFarland in PBS inoculated on Karmali agar without shaking at 42°C for 96 h | • Inconsistent with three independent experiments. | Shagieva et al., 2021 |
| C. coli (AT with AA and AT with non-AA) | Bird | 1 | BA and CCDA | 10 µl drop of 105 CFU on BA without shaking at 37°C for 48 h 10 µl drop of 108 CFU/mL on BA and CCDA without shaking at 42°C for 6 days. |
• Both AA and non-AA strains demonstrated aerobic growth on BA. • Both AA and non-AA strains demonstrated aerobic growth on both BA and CCDA. |
O’Kane and Connerton, 2017 |
| C. jejuni (AT) | Human | 1 | Karmali agar | OD600 = 0.03 in peptone saline solution on Karmali agar without shaking at 42°C for 48 h | • No significant difference in growth rate between C. jejuni prepared in AA and non-AA. | Rodrigues et al., 2015 |
| C. jejuni (AT) | Human, water, and raw chicken | 40 | Nutrient agar | Inoculated from nutrient agar without shaking at 37°C for 5 days | • Majority of isolates grew aerobically. | Chynoweth et al., 1998 |
AT = Aerotolerant; ND = Aerotolerant status not determined, BA=Blood agar; MH = Mueller Hinton; CCDA = Charcoal Cefoperozone Deoxycholate agar; AA = Aerobically acclimatize; PBS = Phosphate Buffered Saline.
Table S6 Summary of three articles on the survival of Campylobacter spp. with different levels of aerotolerance against chemical stress.
| Aerotolerance level of isolates | Isolate origin | Number of isolates | Experimental Media | Incubation conditions | Findings | References |
|---|---|---|---|---|---|---|
| C. jejuni (HAT, AT, and OS) and C. coli (HAT, AT, and OS) | Clinical setting, dairy products, and broiler carcasses | 70 HAT, 25 AT, and 16 OS | Raw chicken skin | Followed Oh et al., 2018 |
• Both HAT and AT strains had significantly better survival than OS strains, with none of the OS strain survived. • Most of the C. jejuni isolates showed significantly greater resistance to PAA than C. coli, with higher final mean cell numbers. |
Mouftah et al., 2021 |
| C. jejuni (HAT, AT, and OS) | Chicken | 25 HAT, 25 AT, and 20 OS | Raw chicken skin | Followed Oh et al., 2018 |
• OS strains had the highest significant reduction in cell number than the AT and HAT strains. | Oh et al., 2019 |
| C. jejuni (HAT, AT, and OS) | Clinical setting | 65 HAT, 46 AT, and 10 OS | Raw chicken skin | OD600 = 0.1 in MH broth, incubated at 4° C under MAC for 1 h for cell attachment. Immerse chicken skin in 750 ppm PAA for 15 s | • Majority of HAT and AT strains survived, while only one OS strain survived. | Oh et al., 2018 |
OS = Aero-sensitive; AT = Aerotolerant; HAT = Hyper-aerotolerant; ND = Aerotolerant status not determined; MH = Mueller Hinton; AC = Aerobic condition; MAC = Microaerobic condition.
Table S7 Summary of thirteen articles on the survival of Campylobacter spp. with different levels of aerotolerance against temperature stress.
| Aerotolerance level of isolates | Isolate origin | Number of isolates | Experimental Media | Incubating conditions | Findings | References |
|---|---|---|---|---|---|---|
| C. jejuni (HAT, AT, and OS) | Retail broiler chicken | 33 HAT, 45 AT, and 12 OS | MH broth | 4°C under AC and MAC conditions for 21 days | • All strains survived under aerobic conditions but not under microaerobic conditions. • Strains viability significantly higher under aerobic conditions. • compared to microaerobic. |
Hur et al., 2024 |
| C. jejuni (HAT, AT, and OS) | Poultry processing plants | 5 HAT, 10 AT, and 25 OS | Broiler chicken drumsticks with skin | 4°C for 7 days (atmosphere not specified) -20°C for 7 days (atmosphere not specified) |
• Significant reduction in HAT compared to OS and AT. • AT survived significantly better than HAT and OS. • HAT survived the least. |
Pokhrel et al., 2023 |
| C. jejuni (HAT, AT, and OS) and C. coli (HAT, AT, and OS) | Clinical strains, dairy products, and broiler carcasses | 70 HAT, 25 AT, and 16 OS | Raw chicken skin | 4°C for 7 days (atmosphere not specified) -20°C for 7 days (atmosphere not specified) |
• Significant reduction in OS and AT strains than HAT strains after 7 days. • More HAT strains survived than AT strains and none of the OS strains survived. |
Mouftah et al., 2021 |
| C. jejuni (HAT, AT, and OS) and C. coli (HAT, AT, and OS) | Clinical strains, dairy products, and broiler carcasses | 70 HAT, 25 AT, and 16 OS | Milk | 72°C for 15 s or 30 s under AC | • Higher proportion of HAT strains survived than AT, and AT strains survived better than OS strains. | Mouftah et al., 2021 |
| C. jejuni (AT and maybe AT*) | Pond water, outlet of wastewater treatment plant, raw chicken meat, and clinical setting | 3 AT and 11 maybe AT | MH broth | 7°C under AC and MAC | • All 14 strains maintained their viability up to 4 weeks under MAC. • Under aerobic incubation, no trend was observed for AT and the other strains. |
Shagieva et al., 2021 |
| C. jejuni (HAT and AT) | Milk, cattle, NCTC 11168, and udder cloth | 6 HAT and 3 AT | Milk | 4°C under AC | • HAT strains survived longer than AT strains. | Jaakkonen et al., 2020 |
| C. coli (HAT and AT) and C. jejuni (OS) | Chicken liver, beef liver, and NCTC 11168 | 1 HAT, 1 AT, and 2 OS | 100% or 10% of chicken meat juice, beef meat juice, beef liver juice, chicken liver juice, 5% laked horse blood and 5% defibrinated horse blood | 4°C under MAC | • Meat juices, liver juices, and horse blood increased the survival of HAT, AT, and OS strains compared to MH broth. | Karki et al., 2019 |
| C. jejuni (AT and OS) | Chicken | 2 AT and 1 OS | BHI | 4°C under AC | • No significant difference in survival between AT and OS. | Lee et al., 2019 |
| C. jejuni (HAT, AT, and OS) | Chicken | 25 HAT, 25 AT, and 20 OS | MH broth | 4°C (not stated atmosphere) -20°C (not stated atmosphere) 72°C for 15 s or 30 s (not stated atmosphere) |
• OS strains exhibit a significantly low survival after storage for seven days compared to HAT and AT. • More HAT and AT strains survived after seven days than OS strains. • 1 HAT and 1 AT strains survived after 30 s. |
Oh et al., 2019 |
| C. jejuni (HAT, AT, and OS) | Clinical setting | 65 HAT, 46 AT, and 10 OS | Chicken skin | 4°C under AC –20°C (not stated atmosphere) |
• Some HAT and AT strains survived after 2 weeks, while only one OS C. jejuni strain survived after 3 days. • Some HAT strains only reduce <1 log10 CFU/g meat after 2 weeks. • Most HAT and AT survived after 7 days. • No OS survived after 3 days. |
Oh et al., 2018 |
| C. jejuni (HAT, AT, and OS) | Clinical setting | 65 HAT, 46 AT, and 10 OS | Whole milk | 72°C for 15 s or 30 s under AC | • Most HAT and AT survived after heat treatments, while no OS strain survived. | Oh et al., 2018 |
| C. jejuni (HAT, AT, and OS) | Chicken | 2 HAT, 2 AT, and 2 OS | Chicken meat and MH broth | 4°C under AC, N2 and CO2 | • In both chicken meat and MH broth under three atmospheric conditions (AC, N2, and CO2), HAT survived longer with a lower cell reduction than AT, while AT survived longer with a lower cell reduction than OS. | Oh et al., 2017 |
| C. jejuni (AT) | NCTC 11351 (bovine) | 1 AT | Autoclaved minced chicken | 5°C under AC | • Both microaerobically-grown and aerobically-grown isolates survived until 19 days, with aerobically-grown isolates had higher survival. | Chynoweth et al., 1998 |
| C. jejuni cocktails (AT) | Human, veterinary case, and raw chicken | 4 AT | Sterilized stream water | 5°C under AC and MAC | • Rate of decline in survival is greater in aerobically-grown C. jejuni cocktail than microaerobically-grown under AC. | Chynoweth et al., 1998 |
| C. jejuni (AT) | 13997 (human) | 1 AT | Blood agar | 5°C under AC and MAC | • For non-aerobic adapted C. jejuni, the strain survived longer under AC than MAC conditions. • Aerobic-adapted C. jejuni survived longer than non-aerobic adapted C. jejuni. |
Jones et al., 1993 |
OS = Aero-sensitive; AT = Aerotolerant; HAT = Hyper-aerotolerant; ND = Aerotolerant status not determined; BA = Blood agar; MH = Mueller Hinton; BHI = Brain heart infusion; AC = Aerobic condition; MAC = Microaerobic condition; *the term “maybe aerotolerant” was stated due to inconsistency in the level of aerotolerance.
Table S8 Summary of three articles on the survival of Campylobacter spp. with different levels of aerotolerance against osmotic stress.
| Aerotolerance level of isolates | Isolate origin | Number of isolates | Experimental Media | Incubating conditions | Findings | References |
|---|---|---|---|---|---|---|
| C. jejuni (HAT, AT, and OS) and C. coli (HAT, AT, and OS) | Clinical setting, dairy products, and broiler carcasses | 70 HAT, 25 AT, and 16 OS | MH broth | OD600 = 0.1 in MH broth with 0, 2, and 4% NaCl in MH broth incubated at 42° C under MAC | • At 4% NaCl, 58% of the HAT strains and 8% of the AT strains survived, while none of the OS strains survived. • At 4% NaCl, the final cell concentration of HAT strains was significantly higher than AT strains while significantly higher in AT than OS strains. |
Mouftah et al., 2021 |
| C. jejuni (HAT, AT, and OS) | Chicken | 25 HAT, 25 AT, and 20 OS | MH agar | OD600 = 0.1 in MH broth, a drop of 10x diluted culture spotted on MH agar supplemented with 1, 2, and 4 % NaCl incubated at 42° C overnight (atmosphere not specified) | • Osmotolerance is not related to aerotolerance. | Oh et al., 2019 |
| C. jejuni (HAT, AT, and OS) | Clinical setting | 65 HAT, 46 AT, and 10 OS | MH agar | 2 and 4 % NaCl incubated at 42°C overnight under MAC | • In 2 % NaCl, higher CFU/mL survived in HAT and AT strains than OS strains. • In 4% NaCl, 38.7% of the HAT and AT strains survived on agar, while none of the OS strains survived. |
Oh et al., 2018 |
OS = Aero-sensitive; AT = Aerotolerant; HAT = Hyper-aerotolerant; MH = Mueller Hinton; AC = Aerobic condition; MAC = Microaerobic condition.
Table S9 Summary of eight articles on the classification of CC based on the level of aerotolerance of C. jejuni and C. coli.
| Strains | Source | Aero-sensitive | Aerotolerant | Hyperaerotolerant | References |
|---|---|---|---|---|---|
| C. jejuni | Clinical setting | – | CC-607 (20%, 1/5) CC-21 (20%, 1/5) ST-904 (CC-607, 20%, 1/5) ST-148 (CC-21, 20%, 1/5) |
CC-354 (20%, 1/5) CC-677 (20%, 1/5) CC-21 (20%, 1/5) ST-354 (CC-354, 20%, 1/5) ST-677 (CC-677, 20%, 1/5) ST-148 (CC-21, 20%, 1/5) |
Ortega-Sanz et al., 2024 |
| C. jejuni | Poultry processing plant | CC-21 (12.5%, 5/40) CC-443 (5%, 2/40) CC-464 (2.5%, 1/40) CC-353 (40%, 16/40) ST-12437 (CC-21, 2.5%, 1/40) ST-8 (CC-21, 10%, 4/40) ST-51 (CC-464, 5%, 2/40) ST-464 (CC-464, 2.5%, 1/40) ST-2132 (CC-353, 15%, 6/40) ST-10382 (CC-353, 2.5%, 1/40) ST-10578 (CC-353, 20%, 8/40) |
CC-443 (7.5%, 3/40) CC-353 (15%, 6/40) CC-UA (2.5%, 1/40) ST-51 (CC-443, 7.5%, 3/40) ST-2132 (CC-353, 5%, 2/40) ST-10578 (CC-353, 10%, 4/40) ST-12439 (CC-UA, 2.5%, 1/40) |
CC-353 (15%, 6/40) ST-2132 (5%, 2/40) ST-10578 (10%, 4/40) |
Pokhrel et al., 2023 |
| C. coli | Swine | CC-828 (85%, 61/72) ND (15%, 11/72) ST-854 (CC-828, 13%, 9/72) ST-1142 (CC-828, 13%, 9/72) ST-1122 (CC-828, 8%, 6/72) ST-1556 (CC-828, 8%, 6/72) ST-8517 (CC-828, 8%, 6/72) ST-887 (CC-828, 7%, 5/72) ST-11645 (ND, 7%, 5/72) ST-2699 (CC-828, 6%, 4/72) ST-1016 (CC-828, 4%, 3/72) ST-1058 (CC-828, 4%, 3/72) ST-1096 (CC-828, 3%, 2/72) ST-1450 (ND, 3%, 2/72) ST-2733 (CC-828, 3%, 2/72) ST-10874 (CC-828, 3%, 2/72) ST-1068 (CC-828, 1%, 1/72) ST-4172 (CC-828, 1%, 1/72) ST-4606 (ND, 1%, 1/72) ST-10668 (ND, 1%, 1/72) ST-10826 (ND, 1%, 1/72) ST-10877 (ND, 1%, 1/72) ST-10879 (CC-828, 1%, 1/72) ST-10928 (CC-828, 1%, 1/72) |
CC-828 (91%, 32/35) ND (9%, 3/35) ST-827 (CC-828, 17%, 6/35) ST-887 (CC-828, 17%, 6/35) ST-1556 (CC-828, 9%, 3/35) ST-2733 (CC-828, 9%, 3/35) ST-854 (CC-828, 6%, 2/35) ST-1122 (CC-828, 6%, 2/35) ST-1142 (CC-828, 6%, 2/35) ST-8517 (CC-828, 6%, 2/35) ST-11645 (ND, 6%, 2/35) ST-830 (CC-828, 3%, 1/35) ST-890 (CC-828, 3%, 1/35) ST-1096 (CC-828, 3%, 1/35) ST-2699 (CC-828, 3%, 1/35) ST-4172 (CC-828, 3%, 1/35) ST-10873 (ND, 3%, 1/35) ST-10927 (CC-828, 3%, 1/35) |
CC-828 (94%, 16/17) ND (6%, 1/17) ST-1068 (CC-828, 18%, 3/17) ST-830 (CC-828, 12%, 2/17) ST-854 (CC-828, 12%, 2/17) ST-827 (CC-828, 6%, 1/17) ST-887 (CC-828, 6%, 1/17) ST-1058 (CC-828, 6%, 1/17) ST-1122 (CC-828, 6%, 1/17) ST-2699 (CC-828, 6%, 1/17) ST-2733 (CC-828, 6%, 1/17) ST-8517 (CC-828, 6%, 1/17) ST-10874 (CC-828, 6%, 1/17) ST-10876 (ND, 6%, 1/17) ST-10879 (CC-828, 6%, 1/17) |
Guk et al., 2021 |
| C. jejuni | Milk Cattle Udder cloth | – | ST-58 (CC-UA, 100%, 3/3) | CC-21 (60%, 3/5) CC-61 (20%, 1/5) CC-45 (20%, 1/5) ST-50 (CC-21, 20%, 1/5) ST-883 (CC-21, 20%, 1/5) ST-43 (CC-21, 20%, 1/5) ST-61 (CC-61, 20%, 1/5) ST-45 (CC-45, 20%, 1/5) |
Jaakkonen et al., 2020 |
| C. coli | Duck | CC-828 (83%, 5/6) CC-UA (17%, 1/6) ST-9575 (CC-828, 33%, 2/6) ST-1860 (CC-828, 17%, 1/6) ST-5507 (CC-828, 17%, 1/6) ST-6148 (CC-UA, 17%, 1/6) ST-7818 (CC-828, 17%, 1/6) |
CC-828 (100%, 22/22) ST-855 (27%, 6/22) ST-829 (27%, 6/22) ST-860 (14%, 6/22) ST-1593 (9%, 2/22) ST-830 (5%, 1/22) ST-832 (5%, 1/22) ST-2711 (5%, 1/22) ST-1586 (5%, 1/22) ST-9867 (5%, 1/22) |
CC-828 (96%, 27/28) CC-UA (4%, 1/28) ST-855 (CC-828, 23%, 6/28) ST-830 (CC-828, 18%, 5/28) ST-829 (CC-828, 14%, 4/28) ST-832 (CC-828, 14%, 4/28) ST-860 (CC-828, 7%, 2/28) ST-828 (CC-828, 4%, 2/28) ST-902 (CC-828, 4%, 2/28) ST-1055 (CC-828, 4%, 1/28) ST-827 (CC-828, 4%, 1/28) ST-1593 (CC-828, 4%, 1/28) ST-6148 (CC-UA, 4%, 1/28) ST-6184 (4%, 1/28) |
Guk et al., 2019 |
| C. jejuni | Human Broiler Cattle | CC-464 (100%, 1/1) | CC-21 (34%, 14/41) CC-460 (24%, 10/41) CC-42 (9%, 4/21) CC-607 (7%, 3/41) CC-464 (7%, 3/41) CC-52 (2%, 1/41) CC-353 (2%, 2/41) CC-22 (2%, 2/41) CC-61 (2%, 1/41) CC-45 (2%, 1/41) |
CC-460 (14%, 4/28) CC-354 (14%, 4/28) CC-52 (14%, 4/28) CC-21 (11%, 3/28) CC-464 (11%, 3/28) CC-607 (7%, 2 /28) CC-353 (7%, 2/41) CC-22 (4%, 1/28) CC-61 (4%, 1/28) CC-48 (4%, 1/28) UA (14%, 4/28) |
Kiatsomphob et al., 2019 |
| C. jejuni | Human | – | CC-403 (100%, 1/1) ST-403 (100%, 1/1) |
- | Rodrigues et al., 2015 |
| C. jejuni | Retail poultry | CC-45 (35%, 7/20) CC-21 (30%, 6/20) CC-48 (5%, 1/20) UA (20%, 4/20) ST-13 (6%, 1/20) ST-21 (6%, 1/20) ST-45 (17%, 3/20) ST-142 (6%, 1/20) ST-659 (6%, 1/20) ST-806 (6%, 1/20) ST-934 (6%, 1/20) ST-1698 (17%, 3/20) ST-1818 (6%, 1/20) ST-3794 (6%, 1/20) ST-4663 (6%, 1/20) ST-4911 (6%, 1/20) ST-6193 (11%, 2/20)UA (11%, 2/20) |
CC-21 (48%, 12/25) CC-45 (12%, 3/25) CC-362 (12%, 3/25) CC-353 (8%, 2/25) CC-354 (4%, 1/25) UA (8%, 2/25) ST-21 (17%, 4/25) ST-43 (4%, 1/25) ST-45 (4%, 1/25) ST-50 (17%, 4/25) ST-137 (4%, 1/25) ST-158 (4%, 1/25) ST-452 (4%, 1/25) ST-587 (13%, 3/25) ST-1086 (4%, 1/25) ST-1698 (4%, 1/25) ST-2359 (4%, 1/25) ST-3484 (4%, 1/25) ST-4681 (4%, 1/25) ST-6193 (4%, 1/25) ST-6261 (4%, 1/25) UA (9%, 2/25) |
CC-21 (48%, 12/25) CC-45 (20%, 5/25) CC-362 (12%, 3/25) CC-353 (4%, 1/25) UA (8%, 2/25) ST-21 (9%, 2/25) ST-43 (4%, 1/25) ST-45 (4%, 1/25) ST-50 (4%, 1/25) ST-137 (4%, 1/25) ST-452 (4%, 1/25) ST-587 (4%, 1/25) ST-806 (4%, 1/25) ST-998 (4%, 1/25) ST-1086 (4%, 1/25) ST-1352 (4%, 1/25) ST-1698 (4%, 1/25) ST-1818 (4%, 1/25) ST-2375 (4%, 1/25) ST-2377 (4%, 1/25) ST-4663 (4%, 1/25) ST-4681 (4%, 1/25) ST-6193 (4%, 1/25) UA (9%, 2/25) |
Oh et al., 2015a |
CC = Clonal complex; ST = Sequence type; UA = Unassigned to any CC; ND = Not determined.
Figure S3 Overview of major findings, research gaps and recommendations of the manuscript.