
Download
ORIGINAL ARTICLE
Exploring the bioactive properties of Cornelian Cherry (Cornus mas L.) fruits and leaves: A pilot study of leaves applications in laying hens’ nutrition
Arabela Elena Untea, Petru Alexandru Vlaicu*, Iulia Varzaru, Alexandra Gabriela Oancea, Mihaela Saracila, Raluca Paula Turcu
Feed and Food Quality Department, National Research and Development Institute for Animal Biology and Nutrition, Balotesti, IF, Romania
Abstract
Cornelian cherry (Cornus mas L.) fruits are well known for their health benefits, but when comparing the -nutritional composition of different parts of the plant, the leaves appear to be a more valuable source of nutrients. The concentrations of carotenoids and vitamin E levels in the leaves were higher than in the fruits. Among individual tocopherols, the most significant difference between fruits and leaves was observed for α-tocopherol. Regarding the polyphenol composition, the major class determined was phenolic acids, expressed as hydroxybenzoic, gallic, and ellagic acids, but the most important was ferulic acid. The major flavonoids quantified were epicatechin and epigallocatechin. Cornelian cherry leaves were selected as a potential feed additive in laying hens’ nutrition, and an experimental trial was performed. Laying hens’ dietary supplementation of Cornelian cherry positively affected egg quality characteristics, by improving the retinol, astaxanthin, and polyphenols concentrations in egg yolk. Tocopherols were negatively influenced, and no significant effect was observed on lutein concentrations. The administration of Cornelian cherry as a dietary supplement produced a favorable impact on oxidative stability during the shelf life of eggs. The Cornelian cherry supplements, rich in natural antioxidants such as polyphenols and vitamins, not only enhance the antioxidant capacity of the animal product but also -contribute to yolk quality maintenance over storage periods by reducing the formation of oxidation products.
Key words: antioxidants, Cornelian cherry, eggs quality, feed additive, laying hen
*Corresponding Author: Petru Alexandru Vlaicu, Feed and Food Quality Department, National Research and Development Institute for Animal Biology and Nutrition, Balotesti, IF, Romania. Email: [email protected]
Academic Editor: Rana Muhammad Aadil, PhD: National Institute of Food Science & Technology, University of Agriculture, Faisalabad, Pakistan
Received: 19 October 2025; Accepted: 25 December 2025; Published: 25 February 2026
© 2026 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
There is a constant concern among nutritionists to -identify new natural sources of bioactive compounds that can maximize the nutritional quality of animal--derived products. For this purpose, unconventional sources of bioactive compounds (plants, coproducts, and by--products that are little or not exploited) and their potential as feed additives in farm animal diets are being evaluated.
The Cornelian cherry (Cornus mas L.), also known as dogwood, is a slow-growing shrub native to all of Europe, except the north, and is adaptable to both temperate and semi-arid regions. The Cornelian cherry is a medicinal and ornamental plant, and its fruits are also valuable sources of bioactive compounds such as vitamin C or polyphenols (Sevindik et al., 2024). The fruits of cornelian cherry are used in traditional European cuisine as cake additives or as raw materials for juices, jams, or liqueurs, with recorded recipes dating back to the 19th century (Kazimierski et al., 2019). Cornelian cherry can be considered an underutilized plant according to its characteristics, such as presenting market value but rarely found for sale, existing in the cultural heritage, and ignored in the food industry (Hossain et al., 2021). In Romania, the dogwood is a wild plant, widespread in hilly and mountainous areas and rarely cultivated as an ornamental plant in parks and gardens (Cosmulescu et al., 2020). Leaves, flowers, seeds, and bark are anatomical parts of Cornelian cherry used in folk medicine, and their therapeutic effects are based on antioxidant, antimicrobial, antidiabetic, antiatherosclerosis, antiobesity properties. Few research studies were focused on the evaluation of nutrients from different plant parts, and they concluded that, along with fruits, other parts such as leaves and flowers are valuable sources of bioactive compounds (Antoniewska-Krzeska et al., 2022). Among the bioactive compounds, polyphenols represent a special class, partly due to their multiple benefits for human health and extensive spread in the plant kingdom. Berries are considered important sources of polyphenolic -compounds, and in the case of colored fruits, anthocyanins are dominant. The major polyphenols in cornelian cherry leaves were reported to be flavonoids (notably quercetin and kaempferol derivatives), phenolic acids (ellagic, chlorogenic, gallic, ferulic, coumaric, caffeic acids), and tannins (Szczepaniak et al., 2019).
The study of the health benefits of berries and berries wastes and their influence on the nutritional quality of animal-based foods is already a topic of major interest discussed in the scientific literature (Vlaicu et al., 2024). Due to competition with human nutrition, animal scientists have discovered rich sources of antioxidants in plant materials with low economic impact, such as leaves, meals or pomaces from the food industry. Studies on the potential use of pomace or leaves of berry trees or shrubs, such as rosehip, cranberries, and blueberries, in poultry feeding have demonstrated beneficial effects on the nutritional quality of poultry products (meat or eggs) (Untea et al., 2019, 2023; Vlaicu et al., 2024), as presented in recent studies.
According to literature data, Cornelian cherry, fruits, leaves or pomaces, represent a novel approach of feed additives for potential utilization poultry nutrition. To date, the available literature regarding the use of dogwood coproducts (fruits and/or leaves) in animal feed or animal-origin foods is very limited. Only a few studies explored these novel sources. In broiler diets, it was reported that using 50, 100, 200, and 400 mg/kg diet of cornelian cherry extract improved the growth performance and positively modulated the intestinal health status. Additionally, the antioxidant capacity and lipid peroxidation of poultry meat were significantly improved without affecting its chemical composition (Ibrahim et al., 2021). The authors suggested that 200 mg/kg is recommended to boost meat shelf stability during long periods of frozen storage. In the production of beef burgers, using 0.5 g or 1.5 g of cornelian cherry juice as a functional additive resulted in beneficial inhibition of oxidative changes and, at the same time, had no negative effect on the sensory characteristics of beef burgers (Salejda et al., 2018). Other authors reported that in the cholesterol-rich diet of rabbits, using 10 or 50 mg/kg of resin-purified cornelian cherry extract, confirms that it may constitute a potentially effective product in the prevention and treatment of obesity-related disorders (Danielewski et al., 2024). However, there is no study exploring the cornelian cherry leaves as a potential source of bioactive compounds, on egg quality, by designing a novel feeding solution for poultry.
In this paper, both the fruits and leaves of C. mas L. were characterized in order to provide a comprehensive overview of the composition and biological value of this species. However, nutritional investigations on egg quality of laying hens have focused exclusively on the leaves, given that the fruits are a valuable resource for human consumption and the food industry, and their use in animal feed would generate undesirable competition with the human food chain. In contrast, the leaves, considered a less exploited by-product, may represent a sustainable alternative resource for integration into animal nutrition, thus contributing to the full utilization of biomass and the reduction of losses.
In this context, the study aimed to provide a comparable overview of the nutritional composition of the fruits and leaves of cornelian cherry. On the other hand, the nutritional potential of Cornelian cherry leaves in laying hens’ diets and its influence on the quality of eggs are also discussed.
Materials and Methods
Chemicals
1,1-Diphenyl-2-picrylhydrazyl (DPPH), disodium ethylenediaminetetracetate (EDTA), trichloroacetic acid (TCA), ferric chloride, thiobarbituric acid (TBA), and ascorbic acid (AA) were purchased from Sigma (Sigma-Aldrich GmbH, Steinheim, Germany). All other chemicals were of analytical grade. Analytical standards were purchased from Sigma-Aldrich (St. Louis, MO, USA). These consisted of δ-tocopherol (95%), γ-tocopherol (96%), and α-tocopherol (96%); lutein (95%), zeaxanthin (95%), and astaxanthin (97%). Phenolic standards: ellagic acid (95%), syringic acid (98%), epicatechin (96%), 4-hydroxy-3-methoxy-cinnamic acid (95%), rutin (95%), vanillic acid (95%), 3-hydroxybenzoic acid (95%), protocatechuic acid (96%), caffeic acid (95%), coumaric acid (98%), epigallocatechin (97%), catechin (95%), quercetin (95%), and resveratrol (99%) were purchased from Sigma (Sigma-Aldrich GmbH, Steinheim, Germany); and ferulic acid (97%) and chlorogenic acid (95%) were purchased from the European Pharmacopoeia (EP).
Vegetal material
The vegetal materials (Cornelian cherry fruits and leaves) were harvested from a hilly area in Dambovita County, Romania (45° 6'4" N 25° 21'29" E). The leaves were collected in the middle of August when they were glossy green in color. The fruits were harvested at the end of September, when they reached the specific shiny red color. The leaves and fruits were dried until they attained constant weight, using a laboratory stove (Memmert UNB 100 Oven, Apeldoorn, Netherlands). The dried products were ground (Grindomix GM 200 mill, Retsch, Haan, Germany) and stored in paper bags.
Plant extracts
Extracts for liposoluble compounds determination
For this extraction, 2 g of plant samples were mixed with 100 mg of butylated hydroxytoluene, 50 mg of ethylenediaminetetraacetic acid, 130 mL of ethanol, 2 mL of sodium ascorbate, and 25 mL of 50% potassium hydroxide solution. The mixture was boiled in a conical flask for 30 min at 80°C in a water bath equipped with a condenser. After boiling, the resulting solution was transferred to a separation funnel and rinsed with 250 mL of water. The conical flask was then washed with 100 mL of petroleum ether and 25 mL of ethanol, which were also transferred to the separation funnel. The solution in the separation funnel was rinsed with water (100 mL). The obtained extract was filtered using anhydrous sodium sulfate. The extract was evaporated in a rotavapor under vacuum conditions, and the resulting residue was dissolved in 10 mL of ethanol (Varzaru et al., 2021).
Extracts for hydrosoluble compounds determination
The preparation method for polyphenols profile determination comprised an extraction step using a solvent mixture of water/methanol/acetic acid (69:30:1, v:v:v) under thermal conditions (50°C) for 1 h. After centrifugation (4500 rpm, 15 min), the supernatant was filtered using a SPE cartridge with silica (1000 mg/6 mL, particle size 40–75 µm, pore size 70 Å). The procedure details were described by Varzaru et al. (2021)
Analytical methods used for cornelian cherry bioactives determination
Liposoluble compounds
The determination of vitamin E isomers was conducted using high-performance liquid chromatography, with a Vanquish Core System (Thermo Fisher Scientific, Bremen, Germany). For the separation, a C18 column, with dimensions: 250 × 4.60 mm, 5 μm (Thermo Fisher Scientific, Bremen, Germany), was used, under isocratic conditions. The mobile phase consisted of methanol (96%) and water (4%), and the flow of the mobile phase was 1 mL/min.
The xanthophylls determination was performed using a Finnigan Surveyor Plus liquid chromatograph (Thermo Fisher Scientific, Bremen, Germany), with a C18 reversed-phase column (250 × 4.60 mm, 5 μm) (Nucleodur, Macherey-Nagel, Duren, Germany). The mobile phase consisted of acetone (75%), methanol (15%), and water (10%), and the flow was set at 1 mL/min. The results of the vitamin E isomers and xanthophylls were reported as milligrams per kilogram of dried samples (mg/kg).
Hydrosoluble compounds
For hydrosoluble compounds like polyphenols, a Vanquish Core HPLC system equipped with a DAD manufactured by Thermo Fisher Scientific (Bremen, Germany) and a BDS HyperSil C 18 column (250 × 4 mm, 5 µm particle size) from Thermo Fisher Scientific (Bremen, Germany) was used. For the separation of polyphenols, a binary gradient, consisting of 1% acetic acid in distilled water (v/v) as solvent A, methanol as solvent B, and acetonitrile as solvent C, with a flow rate set at 0.5 mL/min, was applied. The elution program was as follows: 0–15 min: 5% solvent B, 5% solvent C; 15–20 min: 4% solvent B, 15% solvent C; 20–25 min: 3% solvent B, 25% solvent C; 25–40 min: 2% solvent B, 38% solvent C; and 40–50 min: 5% solvent B, 5% solvent C. The detection wavelengths were 254, 270, 280, 310, and 320 nm. For chromatographic methods, individual standards were used for the identification and quantification of the compounds.
To determine the total polyphenol content, obtaining a methanolic extract was imperative. For extraction, the dried plant sample was mixed with methanol (80%) (1:20). The mixture was placed in a rotary shaker for 24 h in dark conditions. 0.5 mL of the methanolic extract was mixed with 0.5 mL Folin–Ciocâlteu reagent, 2 mL of sodium carbonate solution (20%), and water (7 mL). The resultant solution was kept in the dark for 1 h, and the absorbance was registered at 732 nm using a Jasco V-530 spectrophotometer (Japan Servo Co., Ltd., Japan). The results were reported as mg of gallic acid equivalents per gram of dried sample.
The 2,2-diphenyl-1-picryl-hydrazyl-hydrate method (DPPH) was used for determining the antioxidant capacity. After preparing the DPPH solution in methanol with a 0.2 mM concentration, it was mixed with samples and water in a ratio of 2:0 /4:1/6 (v/v/v). After the mixture was obtained, the absorbance was recorded at 517 nm, using a Jasco V-530 spectrophotometer (Japan Servo Co., Ltd., Japan). The results were expressed as mg of Trolox equivalents per gram of dried sample.
Oxidative stability parameters
Oxidative stability parameters were determined using spectrophotometric methods (V-530 Jasco (Japan Servo Co. Ltd., Japan), described in detail by Untea et al. (2019). Before the determination of the oxidative stability parameters, the extraction of fat was performed using the modified Folch method, with chloroform and methanol as extraction solvents. The samples were mixed in a separation funnel with chloroform 30 mL, methanol 15 mL, and KCl 7.5 mL (0.88%). The lower layer was collected after separation and evaporated under normal conditions. To determine the peroxide value, the ferric thiocyanate method was used. The fat extract was mixed with a solution of chloroform: methanol (7:3), 10 mmol/L xylenol orange, and 1000 mg/kg FeCl2. After obtaining the mixture, it was kept in dark conditions for 5 min. The absorbance was recorded at 560 nm, and the results were reported as milliequivalents of oxygen per kilogram of lipids (meq O2/kg). The conjugated dienes and trienes concentrations were determined using the lipidic extract, mixed with iso-octane. The absorbances were registered at 233 nm for conjugated dienes and 268 nm for conjugated trienes. The p-anisidine value was obtained after registering the absorbance of the lipid extract-iso-octane solution at 350 nm. After that, the p-anisidine reagent was added, and the mixture was placed in the dark for 10 min. A new measurement at 350 nm was performed. For the TBARS values, the samples were mixed with trichloroacetic acid (7.5%) and butylhydroxytoluene (0.8%) in the ratio 2:1. The solutions were centrifuged for 3 min at 3000 g, and filtered. The filtered (2.5 mL) solution was incubated with 1.5 mL of thiobarbituric acid solution (0.8%) in a water bath for 50 min at 80°C, following the recording at 540 nm of the third spectrum.
In vitro induced lipid peroxidation
For the evaluation of the inhibition potential of Cornelian cherry, chicken breast meat samples were used as substrate for peroxidation and FeCl2 (100 µM) and ascorbic acid (500 µM) as oxidants (Untea et al., 2024). After inducing the peroxidation process on fresh meat samples with the oxidant mixture, the methanolic extract was applied. The sample solution was incubated at 37°C for 1 h, and the MDA concentrations were determined according to the TBARS protocol (mixture of sample solution and TCA [7.5%] and BHT [0.8%] react with TBA solution [0.8%] for 50 min at 80°C).
Experimental design
The feeding trial was performed in the National Research and Development Institute for Animal Biology and Nutrition Biobase (Balotesti, Romania), following the Directive 2010/ 63/ EU and Romanian documents 206/ 2004 and 43/ 11.04.2014. The experimental protocol was approved by the Ethical Commission of the National Research and Development Institute for Animal Biology and Nutrition (approval number 3578/17.07.2023). A total of 60 Lohmann Brown Classic laying hens, at 26 weeks old, were randomly allocated to two different pens of 3.96 m2 (0.132 m2/hen), with 30 animals per pen and raised on a litter floor. The feeding trial lasted 8 weeks, two for adaptation, and six for the actual feeding trial. The environmental conditions (humidity, temperature, and ventilation) were monitored using a Viper Touch system. The experimental groups were assigned to the following diets: a control group (CG), which received a conventional diet used in laying hens’ nutrition and an experimental diet with the addition of 0.5% Cornelian cherry leaves (CCG). Both diets were formulated to provide 17.50% crude protein and 2725 kcal/kg metabolizable energy, in accordance with the Lohmann Brown management breeding guide. Water was available ad libitum via nipple drinkers, and each bird received 110 g of feed per day. The lighting regimen was 16 h light and 8 h dark per day. The basal nutritional formula is reported elsewhere (Vlaicu et al., 2024). At the end of the trial, 72 egg samples (36 per group) were collected, and the yolks were analyzed to assess the influence of leaf bioactives on the antioxidant status of the eggs. Eggs collected at the end of the trial were split into two different batches. The first one was immediately analytically evaluated (0 days of storage), and the other was stored at refrigerated temperature (5°C) for 30 days.
Statistical analysis
The results regarding the nutritional composition of C. mas fruits and leaves were expressed as means of triplicate determinations. Statistical differences were assessed by one-way ANOVA followed by Tukey’s HSD test. For the nutritional experimental model, a 2 × 2 factorial arrangement was applied to evaluate the main effects of diet, storage time, and their interaction. Statistical comparisons were performed using two-way ANOVA (General Linear Model procedure), followed by Tukey’s HSD test. Six samples per group were constituted for chemical analysis, each consisting of three egg yolks.
Data related to the evaluation of bioactive compounds from C. mas fruits and leaves, egg yolk nutritional quality and oxidative stability parameters, were processed using XLSTAT software (version 2023.5, Addinsoft, New York, NY, USA). Differences were considered statistically significant at P < 0.05. The lipid peroxidation evaluation, based on an induced oxidation model, was graphically represented using GraphPad Prism version 9.03 (San Diego, CA, USA), with significance set at P < 0.05.
Results and Discussions
The comparative results for liposoluble (carotenoids and vitamins) and hydrosoluble antioxidant (polyphenols) compounds identified in Cornelian cherry fruits and leaves are presented in Tables 1 and 2.
Table 1. Liposoluble bioactive compounds of Cornelian cherry fruits and leaves.
| Items | Fruits | Leaves | SEM | P |
|---|---|---|---|---|
| Carotenoids (mg/kg) | ||||
| Lutein & zeaxanthin | 104.5b | 3988a | 13.19 | 0.0001 |
| Astaxanthin | 0.823b | 2.105a | 0.111 | 0.0001 |
| Canthaxanthin | nd | 1.418 | – | – |
| Beta carotene | 193.5b | 11015a | 23.46 | 0.0001 |
| Vitamins (mg/kg) | ||||
| Alpha tocopherol | 76.37b | 501.3a | 0.893 | 0.0001 |
| Delta tocopherol | 0.102b | 0.158a | 0.006 | 0.003 |
| Gama tocopherol | 11.09b | 21.69a | 0.200 | 0.0001 |
| Tocopherol total | 95.99b | 538.2a | 0.859 | 0.0001 |
SEM, standard error of the mean; P, significance; nd, not determined; letters “a” and “b” on the same row show significant differences at P < 0.05, according to the ANOVA test.
Table 2. Polyphenols profile of Cornelian cherry fruits and leaves.
| Fruits | Leaves | SEM | P | |
|---|---|---|---|---|
| Phenolic acids | ||||
| Hydroxybenzoic acids | ||||
| Gallic acid | 0.961a | 0.325b | 0.006 | 0.0001 |
| Vanillic acid | 0.118 | nd | – | – |
| Syringic acid | 0.360 | nd | – | – |
| Hydroxybenzoic acid | 1.736a | 1.139b | 0.015 | 0.0001 |
| Ellagic acid | 0.745a | 0.009b | 0.004 | 0.0001 |
| Hydroxycinnamic acids | ||||
| Chlorogenic acid | 0.415a | 0.153b | 0.006 | 0.0001 |
| Caffeic acid | 0.177a | 0.016b | 0.005 | 0.0001 |
| Metoxicinnamic acid | nd | 0.017 | – | – |
| Ferulic acid | 2.469a | 0.052b | 0.013 | 0.0001 |
| Coumaric acid | 0.041a | 0.019b | 0.001 | 0.0001 |
| Cinnamic acid | 0.011 | nd | – | – |
| Flavonoids | ||||
| Flavanols | ||||
| Epigallocatechin | 0.901a | 0.039b | 0.003 | 0.0001 |
| Catechin | nd | 0.198 | – | – |
| Epicatechin | 0.707a | 0.011b | 0.004 | 0.0001 |
| Flavonols | ||||
| Rutin | 0.063a | 0.021b | 0.004 | 0.002 |
| Quercetin | nd | nd | – | – |
| Stilbene | ||||
| Resveratrol | 0.023a | 0.004b | 0.001 | 0.0001 |
SEM, standard error of the mean; P, significance; nd, not detected; letters “a” and “b” on the same row show significant differences at P < 0.05, according to the ANOVA test.
Cornelian cherry fruits are well known for their health benefits, both in scientific literature and traditional medicine. However, when comparing the nutritional composition of different parts of the plant, the leaves appear to be a more valuable source of nutrients. For all analyzed carotenoids, concentrations in the leaves were higher than in the fruits, ranging from 2.5 to over 50 times greater. Similarly, vitamin E levels in the leaves were up to five times higher. Among individual tocopherols, the most significant difference between fruits and leaves was observed for alpha-tocopherol (Table 1).
The fruits of C. mas are an exceptional source of vitamin C (Lidiková et al., 2024), but the leaves proved to be a rich source of liposoluble antioxidants (vitamin E and carotenoids) and present potential use in functional foods and feeds. Other researchers mentioned the existence of tocopherol, biotin, and riboflavin in aqueous extracts of C. mas (Tenuta et al., 2022), but did not report numerical values. Scientific papers that followed the comparative content (fruits vs leaves) of vitamins and carotenoids from plants belonging to the “berry” class reported important concentrations in leaves compared to fruits. A recent study on black elderberry showed that vitamin E content in leaves ranged from 726.12 to 1486.58 mg/kg, while the fruits ranged from 51.75 to 61.17 mg/kg (Vlaicu et al., 2025). Although the fruits of Cornelian cherry contain higher vitamin E content than black elderberry, the leaves contain scarce amounts of this bioactive compound. The same authors reported that fruits are the main source of carotenoids in black elderberry, especially lutein and -zeaxanthin (4782.85 mg/kg), while in Cornelian cherry, the leaves are the major source of this compound (288.76 mg/kg). In line with our results, another comparative study on black chokeberry found leaves to be the main deposits of liposoluble compounds, vitamin E (1172.20 mg/kg), and xanthophylls (2353.24 mg/kg) (Saracila et al., 2024). These differentiated results among different plants and their coproducts have been attributed to the defense system of plants against stressors; in some cases, more antioxidants are concentrated in the leaves compared to fruits for plant protection (Vlaicu et al., 2025). The study results and the differentiated deposition of bioactive compounds in different plant parts make leaves a subject of further research.
Cornelian cherry is considered a valuable source of polyphenols (Table 2), with fruits rich in anthocyanins and iridoids and leaves containing phenolic acids and flavonoids (Antoniewska-Krzeska et al., 2024). In terms of total polyphenol content, the leaves are a rich source of these compounds, with their concentrations reaching values up to 10 times higher than those of the fruits in some cases (Bayram et al., 2020). Badalica-Petrescu et al. (2014) identified 15 different phenolic molecules (aglycons) using HPLC-DAD-ESI (+) MS analysis. The major categories of phenolics identified were epicatechin, derivatives of coumaric acid, caffeic acid, quercetin, and kaempferol. Our results revealed phenolic acids as major components, expressed as hydroxybenzoic, gallic, and ellagic acids, but the most important value was determined for ferulic acid. Similarly, Cosmulescu et al. (2018) reported the highest average values for gallic and coumaric acids (14.49 mg/100g), ellagic acid (5.71 mg/100g), and ferulic acid (1.25 mg/100g) in cornelian fruits. The major flavonoids quantified were epicatechin and epigallocatechin. These substances contribute to the nutritional and functional value of the fruit for human consumption. In terms of leaves, when compared with other leaves belonging to the berry family, such as blackberry or raspberry (Varzaru et al., 2023), with the same extraction, separation, and quantification method, Cornelian cherry proved to be an important source of ferulic acid, while the other berry leaves (blackcurrant, cranberry and aronia leaves extracts) were abundant in chlorogenic acid or epicatechin (Saracila et al., 2024; Untea et al., 2023, 2024). These observations are in line with those recently reported by Forman et al. (2025), where leaves of nine C. species were analyzed. The authors showed that chlorogenic acid ranged from 0.43 to 18.41 mg/g, gallic from 0.26 to 1.63 mg/g, and ellagic acid values from 0.03 to 1.17 mg/g. These results show that the same plant species can result in different polyphenol compositions; however, all of them provide a significant total of polyphenol content, with benefits in poultry nutrition and egg quality.
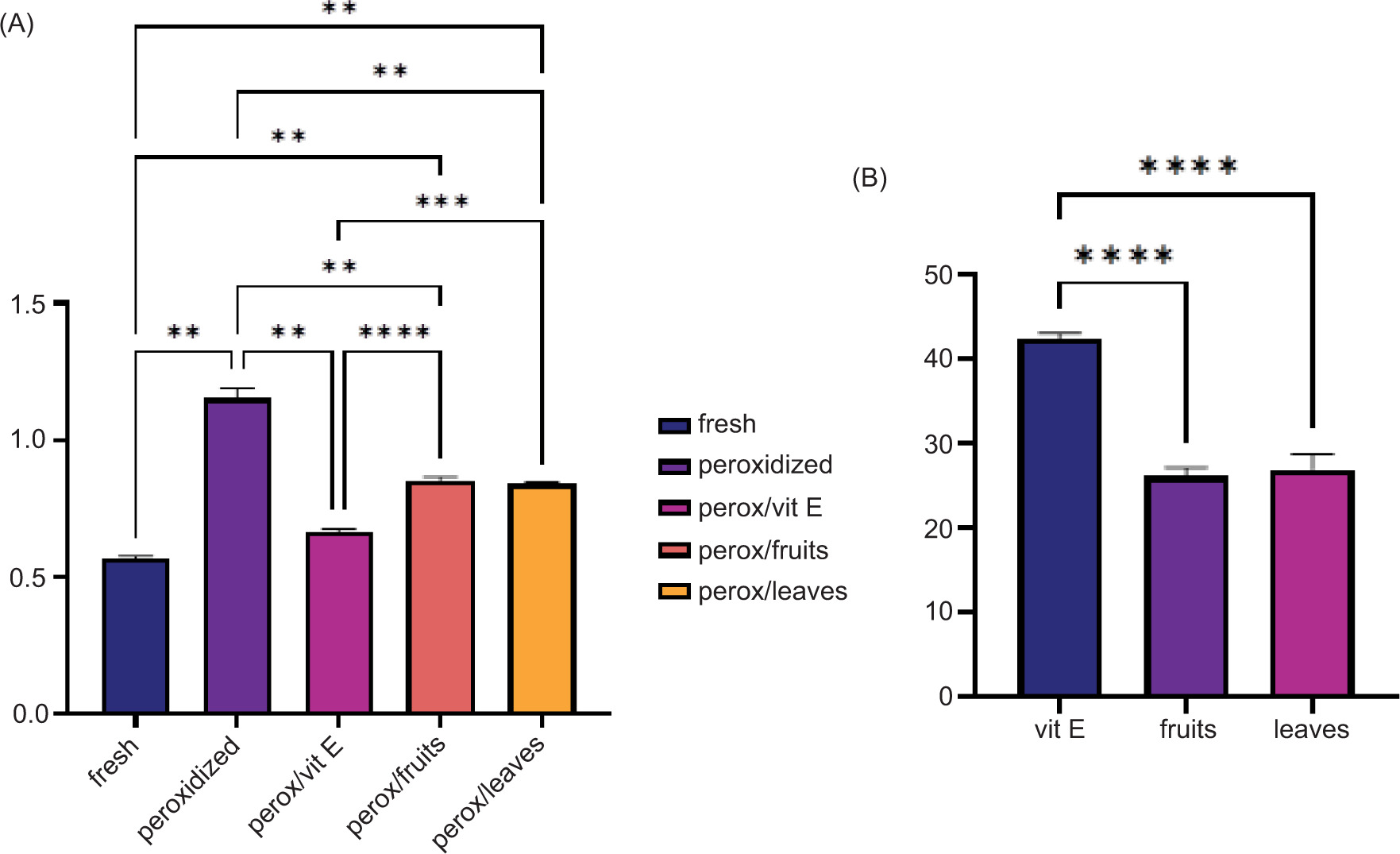
In vitro studies evaluating the lipid peroxidation inhibition potential of plant extracts typically involve a biological model system composed of a lipid-rich animal matrix, an oxidative catalyst, most commonly a transition metal ion such as Fe2+ to induce peroxidation and the plant extract under investigation as the inhibitory agent. This setup allows for the assessment of the extract’s efficacy in reducing the formation of lipid peroxidation markers measured as malondialdehyde (MDA) products formed (Figure 1). The primary mechanism by which iron induces lipid peroxidation is the Fenton reaction, where ferrous iron catalyzes the production of reactive hydroxyl radicals and initiates the free radical chain reactions that degrade polyunsaturated fatty acids in meat lipids (Yadav et al., 2023). In practice, iron and ascorbic acid are commonly used together in meat models to induce lipid peroxidation. Ascorbic acid reduces ferric iron (Fe3+) to ferrous iron (Fe2+), increasing the availability of reactive ferrous iron that participates in the Fenton reaction, and this synergistic action accelerates the oxidation of lipids in the meat model. The data presented in Figure 1 indicate that the MDA concentration in fresh meat was 0.568 ± 0.018 mg/kg, whereas in the peroxidized meat, this value nearly doubled, reaching 1.151 ± 0.075 mg/kg, confirming the oxidative impact of the iron/ascorbic acid pro-oxidant system. Treatment with a synthetic antioxidant (vitamin E) led to a significant reduction in lipid peroxidation, with MDA levels decreasing to 0.663 ± 0.028 mg/kg. The addition of C. mas fruit and leaf extracts also showed antioxidant activity, though to a lesser extent than vitamin E, resulting in MDA concentrations of 0.850 ± 0.031 mg/kg and 0.839 ± 0.014 mg/kg, respectively. No statistically significant difference was observed between the effects of the two plant extracts. The inhibition percentage of Cornelian cherry extracts was 25.96 ± 2.19 for fruits and 26.85 ± 3.63 for leaves, with no statistical differences between them. The extract’s inhibition capacities were significantly decreased compared with the vitamin E potential (42.31 ± 1.32%). The in vitro inhibition activity of extracts proved their antioxidant potential in delaying lipid peroxidation processes. Several plant extracts (Indian spices and Acacia, olive leaves, blackcurrant [fruits, pomace, and leaves], blackberry, and raspberry leaves) have been reported in the scientific literature to be very efficient in counteracting the degradation processes by applying in vitro peroxidation models, and shown to inhibit iron-induced lipid peroxidation, largely due to their antioxidant and iron-chelating properties (Botsoglou et al., 2013; Untea et al., 2024; Varzaru et al., 2023; Yadav et al., 2023).
Figure 1. In vitro induced lipid peroxidation on lipid-rich animal matrix (meat). The peroxidation of meat measured as MDA mg/kg under vitamin E and Cornelian cherry extracts (A) and the inhibitory effect of vitamin E and Cornelian cherry fruits and leaves extract (B). level of significance **P < 0.001, ***P < 0.0001, ****P < 0.00001.
The leaves of Cornelian cherry proved to be a valuable source of bioactive compounds with potential in animal nutrition. To evaluate their effect on laying hens, a diet supplemented with 0.5% Cornelian cherry leaf powder was formulated. To our knowledge, no scientific data has been reported on the inclusion of cornelian cherry leaves in the nutrition of laying hens. An experimental feeding trial was conducted, and the results indicated that the inclusion of the plant material had no significant impact on productive performance parameters compared to the control group (data not shown). Liposoluble bioactive compounds of egg yolk from hens fed with a conventional diet, compared with 0.5% Cornelian cherry leaves-supplemented diet (0 vs 30 days storage time), are presented in Tables 3 and 4.
Table 3. Vitamin concentrations in yolk (0 vs 30 days storage time).
| Time | Diet | α tocopherol, mg/kg |
δ tocopherol, mg/kg |
γ tocopherol, mg/kg |
Tocopherol total, mg/kg |
Retinol, mg/kg |
|---|---|---|---|---|---|---|
| 0 days | CG | 205.2a | 3.869a | 17.19a | 226.2a | 24.48a |
| CCG | 157.2c | 3.618ab | 16.16a | 177.0c | 25.78a | |
| 30 days | CG | 186.8b | 3.696ab | 13.37b | 203.8b | 19.18b |
| CCG | 148.5c | 3.522b | 11.67b | 163.7c | 21.18b | |
| Main effects | ||||||
| Diet | CG | 195.9a | 3.783a | 15.28a | 215.0a | 21.83b |
| CCG | 152.8b | 3.570b | 13.92b | 170.4b | 23.48a | |
| Time | 0 days | 181.2a | 3.743 | 16.67a | 201.6a | 25.14a |
| 30 days | 167.6b | 3.609 | 12.52b | 183.7b | 20.18b | |
| P | ||||||
| Diet | 0.001 | 0.004 | 0.378 | 0.001 | 0.580 | |
| Time | 0.001 | 0.053 | 0.378 | 0.001 | 0.580 | |
| Diet × Time | 0.110 | 0.560 | 0.534 | 0.195 | 0.821 | |
| SEM | ||||||
| Diet | 2.041 | 0.046 | 0.019 | 2.394 | 0.048 | |
| Time | 2.041 | 0.046 | 0.001 | 2.394 | 0.001 | |
| Diet × Time | 2.887 | 0.065 | 0.538 | 3.386 | 0.678 | |
SEM, standard error of the mean; P, significance; letters “a” and “b” on the same row show significant differences at P < 0.05, according to the ANOVA test.
Table 4. Xanthophyll concentrations in yolk (0 vs 30 days storage time).
| Time | Diet | Lutein (mg/kg) |
Astaxanthin (mg/kg) |
Canthaxanthin (mg/kg) |
|---|---|---|---|---|
| 0 days | CG | 33.51ab | 10.23b | 11.32a |
| CCG | 34.35a | 14.49a | 10.80ab | |
| 30 days | CG | 31.27bc | 5.935c | 9.006b |
| CCG | 29.40c | 5.088c | 8.851b | |
| Main effects | ||||
| Diet | CG | 31.87 | 8.083b | 10.16 |
| CCG | 32.38 | 9.791a | 9.828 | |
| Time | 0 days | 33.93a | 12.36a | 11.06a |
| 30 days | 30.33b | 5.512b | 8.929b | |
| P | ||||
| Diet | 0.481 | 0.006 | 0.537 | |
| Time | 0.001 | 0.001 | 0.002 | |
| Diet × Time | 0.080 | 0.001 | 0.739 | |
| SEM | ||||
| Diet | 0.500 | 0.360 | ||
| Time | 0.500 | 0.360 | 0.374 | |
| Diet × Time | 0.707 | 0.510 | 0.529 | |
SEM, standard error of the mean; P, significance; letters “a” and “b” on the same row show significant differences at P < 0.05, according to the ANOVA test.
Data shown in Table 3 presented the decreasing effects of dietary supplements on the tocopherol levels in the yolk. Only vitamin A registered significant increased concentration in the experimental group, compared with the control. To date, there is no evidence regarding the effect of cornelian cherry supplementation on the vitamin E content of egg yolk in laying hens. However, other studies on the effect of dietary vitamin E or phytogenics on tocopherol content of egg yolk prove a direct relationship between the antioxidants from diets and their concentrations in the end products (eggs) (Rbah et al., 2025; Zhao et al., 2021). The mechanism that can explain the influence of dietary antioxidants on egg yolk nutrient content is centered on diet-driven maternal transfer and antioxidant interactions. Liposoluble compounds are absorbed from the hen’s diet, transported via the bloodstream bound to lipoproteins, and deposited in the yolk during its formation (Surai et al., 2001). Other studies proved that carotenoids and vitamin E deposition in yolk are diet-dependent and influenced by environmental factors, and a competitive absorption in nutrient uptake is present. A nutrition study on humans proved that lutein absorption is not influenced by the presence of vitamin E in the diet but can be impaired by other carotenoids and the polyphenol content (Reboul et al., 2007). Other researchers also found that vitamin E competes for absorption with some carotenoids, the same site of absorption being a possible explanation (Saracila et al., 2022). In broiler breeder hens nutrition, high vitamin A content (20,000–35,000 IU/kg) had significant detrimental effects, leading to decreased liver function, reproductive performance, and immune response (Yuan et al., 2014). The authors showed that increasing the content of vitamin A (5000–35,000 IU/kg) led to progressive increase of this bioactive compound in eggs and liver, however, vitamin E and isomers, registered progressive decrease from 15.68 to 8.58 mg/100 g egg yolk. Based on these findings, we assumed that due to the high content of β-carotene (11,015 mg/kg) in the cornelian cherry leaves, vitamin E deposition in the egg yolks was decreased.
Regarding the xanthophyll levels (Table 4), only astaxanthin was positively influenced by the dietary supplements in fresh eggs. The time was an influencing factor on the nutritional quality xanthophyll in eggs. Decreasing values were determined after 30 days of storage in refrigerator (5°C). Previous studies reported that xanthophylls with high polarity (lutein), registered moderate decreases (up to 10–20%) after 30 days of refrigerated storage, attributed to their susceptibility to oxidation and isomerization in the lipid matrix of the egg (Dansou et al., 2023; Dias et al., 2014). In contrast, astaxanthin, although considered more structurally stable, initially presented significantly higher values in the group supplemented with cornelian cherry leaves, decreased proportionally during storage. The results align with previous observations that xanthophyll carotenoids (lutein) are more stable in the long term than ketocarotenoids (astaxanthin), and plant supplementation alters their initial distribution in the eggs during storage, as recently reported (Chen et al., 2025). The competition between liposoluble compounds for absorption cannot explain the decreased deposition of vitamins and xanthophylls in yolks belonging to C. mas supplemented fed group. This reduction can be attributed to the preferential consumption of astaxanthin as an antioxidant for the protection of yolk lipids, a phenomenon also described in other studies on the stability of carotenoids in enriched eggs (Chen et al., 2025). Another potential explanation is the antinutritional factors present in Cornelian Cherry leaves that may inhibit liposoluble compounds absorption and deposition. Although C. mas has potential as a natural antioxidant source, tannins, phytic acid, high fiber, or saponins may affect the metabolism of carotenoids (Chukwuebuka et al., 2015). A study regarding the physicochemical properties of cornelian cherry revealed that the hydrolysable tannins are the major class of phenolics in the fruit matrix of the studied plant (Güzel et al., 2021). Other authors cited in the study noticed remarkable tannin concentrations in C. mas, which ranged between 0.6 and 14% (Dinda et al., 2016), which might also contribute to the xanthophyll’s decrease during storage.
Table 5 presents comparative data regarding the effect of cornelian cherry supplementation in diets of laying hens on polyphenols and DPPH values recorded in egg yolks at the end of the experiment and 0after 30 days of storage at refrigeration temperature. The results obtained revealed a significant effect of supplements on concentrations of total polyphenols and DPPH, and the antioxidant status of eggs was not affected by the storage time. In human nutrition, vegetal sources are recognized as the primary providers of hydrosoluble antioxidants, such as polyphenols. The chemical composition of egg yolk typically reveals low concentrations of polyphenols compared to the plant kingdom. However, some studies in animal nutrition have demonstrated the potential of phytoadditives in increasing polyphenol concentrations in egg yolk and enhancing the total antioxidant activity of the product. For instance, vegetable by-products from the food industry, such as grape pomace (Herranz et al., 2024), pomegranate peel (Lioliopoulou et al., 2024), and rosehip meal (Vlaicu et al., 2022), as well as phytogenic additives such as bilberry and walnut leaves (Untea et al., 2020), have been identified as effective nutritional vectors capable of establishing a functional link between dietary supplementation and egg yolk enrichment. The results obtained in the present study found that the supplementation of diets with cornelian cherry leaves led to increased polyphenol concentrations in the yolks of laying hens. This was associated with an improved antioxidant activity, which remained stable throughout storage, suggesting a positive impact on the oxidative stability of the product.
Table 5. Total polyphenols and antioxidant capacity determined in yolk (0 vs 30 days storage time).
| Time | Diet | Total polyphenols (mg/g GAE) | DPPH (mg/g Trolox) |
|---|---|---|---|
| 0 days | CG | 0.666b | 1.363ab |
| CCG | 0.820a | 1.624a | |
| 30 days | CG | 0.602b | 1.220b |
| CCG | 0.641b | 1.524a | |
| Main effects | |||
| Diet | CG | 0.634b | 1.292b |
| CCG | 0.730a | 1.574a | |
| Time | 0 days | 0.742a | 1.494 |
| 30 days | 0.621b | 1.372 | |
| P | |||
| Diet | 0.009 | 0.001 | |
| Time | 0.002 | 0.102 | |
| Diet × Time | 0.092 | 0.765 | |
| SEM | |||
| Diet | 0.023 | 0.050 | |
| Time | 0.021 | 0.050 | |
| Diet × Time | 0.030 | 0.070 | |
SEM, standard error of the mean; P, significance; nd, not detected; letters “a” and “b” on the same row show significant differences at P < 0.05, according to the ANOVA test.
Oxidative indices were assessed at the end of the experimental period and after 30 days of refrigerated storage (Table 6). At the initial evaluation point, no significant differences were observed between the experimental groups for any of the analyzed parameters. The overall effect showed a dietary positive influence on peroxide value and TBARS, and the time effect was present on conjugated dienes and trienes, p-anisidine value and TBARS. The results presented in the previous tables showed a positive influence of the administered supplements on certain nutrients with antioxidant potential, such as polyphenols and some carotenoids. The presence of bioactive compounds with antioxidant potential, found even at the level of the egg yolk, led to increased oxidative stability during the storage period. This can be a possible explanation for the effect observed (directly transferring antioxidant compounds into the egg yolk), and/or another possible explanation could be enhancing the animal’s endogenous antioxidant defense system. Other authors have also reported positive effects of phytogenic additives on the oxidative stability of egg yolk. Thus, the use of bilberry or walnut leaves (Untea et al., 2020), rosehip meal (Vlaicu et al., 2022), grape pomace (Romero et al., 2022), or pomegranate peel (Lioliopoulou et al., 2023) as supplements in laying hens’ diets has led to improved yolk antioxidant parameters and delayed lipid oxidation during storage.
Table 6. Oxidative stability parameters determined in yolk (0 vs 30 days storage time).
| Time | Diet | Peroxide value (meq active O2/kg) |
Conjugated dienes (µmol/g) |
Conjugated trienes (µmol/g) |
P-anisidine value | TBARS (µg/g) |
|---|---|---|---|---|---|---|
| 0 days | CG | 0.219ab | 6.742 | 2.382 | 15.09b | 136.4b |
| CCG | 0.135b | 6.560 | 2.678 | 9.227b | 116.6b | |
| 30 days | CG | 0.268a | 7.414 | 2.943 | 31.02a | 214.2a |
| CCG | 0.160b | 7.400 | 3.179 | 27.40a | 166.0ab | |
| Main effects | ||||||
| Diet | CG | 0.243a | 6.980 | 2.662 | 23.05 | 175.3a |
| CCG | 0.148b | 7.078 | 2.928 | 18.31 | 141.3b | |
| Time | 0 days | 0.210 | 6.651b | 2.530b | 12.16b | 126.5b |
| 30 days | 0.247 | 7.407a | 3.061a | 29.21a | 190.1a | |
| P | ||||||
| Diet | 0.001 | 0.786 | 0.229 | 0.059 | 0.022 | |
| Time | 0.109 | 0.046 | 0.022 | 0.001 | 0.0001 | |
| Diet x Time | 0.595 | 0.817 | 0.891 | 0.642 | 0.310 | |
| SEM | ||||||
| Diet | 0.016 | 0.252 | 0.152 | 1.678 | 9.691 | |
| Time | 0.016 | 0.252 | 0.152 | 1.678 | 9.691 | |
| Diet x Time | 0.022 | 0.356 | 0.214 | 2.373 | 13.70 | |
SEM, standard error of the mean; P, significance; letters “a” and “b” on the same row show significant differences at P < 0.05, according to the ANOVA test.
Conclusions
Cornelian cherry is a valuable source of antioxidant compounds like vitamins, xanthophylls, or polyphenols. Among different parts of the plant, the obtained results showed that leaves are the main deposit of bioactive compounds. The use of Cornelian cherry in laying hens’ diets, can be a solution to increase the nutritional quality of egg yolks with favorable impact on oxidative stability during shelf life of eggs. Research studies on the possibilities of including Cornelian cherry leaves in the diet of laying hens will continue, as further data is needed to complete the current knowledge.
Data Availability Statement
The data is contained within the article.
Mandatory Disclosure on Use of Artificial Intelligence
The authors declare that no AI-assisted tools were used in the preparation of this manuscript. All references have been manually verified for accuracy and relevance.
Author Contributions
Conceptualization, A.E.U and P.A.V.; methodology, A.E.U., P.A.V.; I.V., A.G.O., M.S., R.P.T.; formal analysis, A.E.U.; P.A.V.; I.V., A.G.O., M.S., R.P.T., writing—original draft preparation, A.E.U.; writing—review and editing, A.E.U and P.A.V.; project administration, P.A.V. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Funding
This work was supported by a grant from the Ministry of Education and Research, CNCS – UEFISCDI, Project OXICAP, Grant number 11TE/2025, within PNCDI IV.
REFERENCES
Antoniewska-Krzeska, A., Ivanišová, E., Klymenko, S., Bieniek, A.A., Šramková, K.F., Brindza, J., 2022. Nutrients content and composition in different morphological parts of Cornelian cherry (Cornus mas L.). Agrobiodiversity for Improving Nutrition, Health and Life Quality 6: 1. 10.15414/ainhlq.2022.0001
Badalica-Petrescu, M., Dragan, S., Ranga, F., Fetea, F., Socaciu, C., 2014. Comparative HPLC–DAD–ESI(+)MS fingerprint and quantification of phenolic and flavonoid composition of aqueous leaf extracts of Cornus mas and Crataegus monogyna, in relation to their cardiotonic potential. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 42: 9–18. 10.15835/nbha4219270
Bayram, H.M., and Ozturkcan, S.A., 2020. Bioactive components and biological properties of cornelian cherry (Cornus mas L.): A comprehensive review. Journal of Functional Foods 75: 104252. 10.1016/j.jff.2020.104252
Botsoglou, E.N., Govaris, A.K., Ambrosiadis, I.A., and Fletouris, D.J., 2013. Olive leaves (Olea europaea L.) versus α-tocopheryl acetate as dietary supplements for enhancing the oxidative stability of eggs enriched with very-long-chain n-3 fatty acids. Journal of the Science of Food and Agriculture 93: 2053–2060. 10.1002/jsfa.6017
Chen, X., Yan, Z., Zhang, B., Zeng, L., Chowdhury, U., Pabitra, M.H., et al. 2025. Lutein and astaxanthin supplementation induce competitive inhibition of carotenoid deposition in egg yolk. Animals 15: 1869. 10.3390/ani15131869
Chukwuebuka, E., and Chinenye, I.J., 2015. Biological functions and anti-nutritional effects of phytochemicals in living system. Journal of Pharmaceutical and Biological Sciences 10: 10–19. 10.9790/3008-10231019
Cosmulescu, S., Cornescu Fratutu, F., and Radutoiu, D., 2020. Determination of morphological characteristics of leaves in cornelian cherry (Cornus mas L.). Romanian Biotechnological Letters 25: 1754–1758.
Cosmulescu, S.N., Trandafir, I., and Cornescu, F., 2018. Antioxidant capacity, total phenols, total flavonoids and colour component of Cornelian cherry (Cornus mas L.) wild genotypes. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 47(2): 390–394. 10.15835/nbha47111375
Danielewski, M., Rapak, A., Kruszyńska, A., Małodobra-Mazur, M., Oleszkiewicz, P., Dzimira, S., et al. 2024. Cornelian cherry (Cornus mas L.) fruit extract lowers SREBP-1c and C/EBPα in liver and alters various PPAR-α, PPAR-γ, LXR-α target genes in cholesterol-rich diet rabbit model. International Journal of Molecular Sciences 25: 1199. 10.3390/ijms25021199
Dansou, D.M., Zhang, H., Yu, Y., Wang, H., Tang, C., Zhao, Q., et al. 2023. Carotenoid enrichment in eggs: From biochemistry perspective. Animal Nutrition 14: 315–333. 10.1016/j.aninu.2023.05.012
Dias, M.G., Camões, M.F.G., and Oliveira, L., 2014. Carotenoid stability in fruits, vegetables and working standards–Effect of storage temperature and time. Food Chemistry 156: 37–41. 10.1016/j.foodchem.2014.01.050
Dinda, B., Kyriakopoulos, A.M., Dinda, S., Zoumpourlis, V., Thomaidis, N.S., Velegraki, A., et al. 2016. Cornus mas L. (cornelian cherry), an important European and Asian traditional food and medicine: Ethnomedicine, phytochemistry and pharmacology for its commercial utilization in drug industry. Journal of Ethnopharmacology 193: 670–690. 10.1016/j.jep.2016.09.042
Forman, V., Trush, K., Konôpková, J., Ferus, P., Czigle, S., Mučaji, P., et al. 2025. Phenolic compounds profiling of nine dogwood species (Cornus L.) leaves. PeerJ 13: e19457. 10.7717/peerj.19457
Güzel, N., 2021. Morphometric and physico-chemical properties of Cornelian cherry (Cornus mas L.) grown in Çorum, Turkey. Akademik Gıda 19: 373–380. 10.24323/akademik-gida.1050750
Halliwell, B., and Gutteridge, J.M.C., 2015. Free radicals in biology and medicine. 5th ed. Oxford, UK: Oxford University Press. 10.1093/acprof:oso/9780198717478.001.0001
Herranz, B., Romero, C., Sánchez-Román, I., López-Torres, M., Viveros, A., Arija, I., et al. 2024. Enriching eggs with bioactive compounds through the inclusion of grape pomace in laying hens diet: Effect on internal and external egg quality parameters. Foods 13: 1553. 10.3390/foods13101553
Hossain, M.M., Rahim, M.A., and Haque, M.R., 2021. Biochemical properties of some important underutilized minor fruits. Journal of Agriculture Food Research 5: 100148. 10.1016/j.jafr.2021.100148
Ibrahim, D., Moustafa, A., Metwally, A.S., Nassan, M.A., Abdallah, K., Eldemery, F., et al. 2021. Potential application of Cornelian cherry extract on broiler chickens: Growth, expression of antioxidant biomarker and glucose transport genes, and oxidative stability of frozen meat. Animals 11: 1038. 10.3390/ani11041038
Kazimierski, M., Reguła, J., and Molska, M., 2019. Cornelian cherry (Cornus mas L.)—Characteristics, nutritional and pro-health properties. Acta Scientiarum Polonorum, Technologia Alimentaria 18: 5–12. 10.17306/J.AFS.2019.0628
Lidiková, J., Čeryová, N., Grygorieva, O., Bobková, A., Bobko, M., Árvay, J., et al. 2024. Cornelian cherry (Cornus mas L.) as a promising source of antioxidant phenolic substances and minerals. European Food Research and Technology 250: 1745–1754. 10.1007/s00217-024-04513-z
Lioliopoulou, S., Papadopoulos, G.A., Giannenas, I., Vasilopoulou, K., Squires, C., Fortomaris, P., et al. 2023. Effects of dietary supplementation of pomegranate peel with xylanase on egg quality and antioxidant parameters in laying hens. Antioxidants 12: 208. 10.3390/antiox12010208
Lioliopoulou, S., Papadopoulos, G.A., Mantzouridou, F.T., Giannenas, I., Kalogeropoulou, A.G., Lioupi, A., et al. 2024. Pomegranate peel dietary supplementation in laying hens affects egg yolk color and eggshell quality, decreases yolk lipid oxidation and influences yolk lipidomic profile. Journal of Applied Poultry Research 33: 100495. 10.1016/j.japr.2024.100495
Rbah, Y., Taaifi, Y., Allay, A., Mansouri, F., Belhaj, K., Houmy, N., et al. 2025. Optimization of hemp seed supplementation with phytobiotics in laying hen feed to improve egg yolk fatty acids, tocopherols and cholesterol using response surface models. Frontiers in Sustainable Food Systems 9: 1566447. 10.3389/fsufs.2025.1566447
Reboul, E., Thap, S., Tourniaire, F., André, M., Juhel, C., Morange, S., et al. 2007. Differential effect of dietary antioxidant classes (carotenoids, polyphenols, vitamins C and E) on lutein absorption. British Journal of Nutrition 97: 440–446. 10.1017/S0007114507352604
Romero, C., Arija, I., Viveros, A., and Chamorro, S. 2022. Productive performance, egg quality and yolk lipid oxidation in laying hens fed diets including grape pomace or grape extract. Animals 12: 1076. 10.3390/ani12091076
Salejda, A.M., Kucharska, A.Z., and Krasnowska, G., 2018. Effect of Cornelian cherry (Cornus mas L.) juice on selected quality properties of beef burgers. Journal of Food Quality 1: 1563651. 10.1155/2018/1563651
Saracila, M., Untea, A.E., Oancea, A.G., Varzaru, I., and Vlaicu, P.A., 2024. Comparative analysis of black chokeberry (Aronia melanocarpa L.) fruit, leaves, and pomace for their phytochemical composition, antioxidant potential, and polyphenol bioaccessibility. Foods 13: 1856. 10.3390/foods13121856
Saracila, M., Untea, A.E., Panaite, T.D., Varzaru, I., Oancea, A., Turcu, R.P., et al. 2022. Creeping wood sorrel and chromium picolinate effect on the nutritional composition and lipid oxidative stability of broiler meat. Antioxidants 11: 780. 10.3390/antiox11040780
Sevindik, M., Khassanov, V.T., Sevindik, E., Uysal, I., and Mohammed, F.S., 2024. Cornelian Cherry (Cornus mas L.): A comprehensive review on its usage areas, biological activities, mineral, phenolic and chemical contents and applications. Applied Fruit Science 66(5): 2061–2071. 10.1007/s10341-024-01151-3
Surai, P.F., Speake, B.K., and Sparks, N.H.C., 2001. Carotenoids in avian nutrition and embryonic development. Absorption, availability and levels in plasma and egg yolk. The Journal of Poultry Science 38: 1–27. 10.2141/jpsa.38.1
Szczepaniak, O.M., Kobus-Cisowska, J., Kusek, W., and Przeor, M., 2019. Functional properties of Cornelian cherry (Cornus mas L.): A comprehensive review. European Food Research and Technology 245: 2071–2087. 10.1007/s00217-019-03313-0
Tenuta, M.C., Deguin, B., Loizzo, M.R., Cuyamendous, C., Bonesi, M., Sicari, V., et al. 2022. An overview of traditional uses, phytochemical compositions and biological activities of edible fruits of European and Asian Cornus species. Foods 11: 1240. 10.3390/foods11091240
Untea, A.E., Oancea, A.-G., Vlaicu, P.A., Varzaru, I., and Saracila, M. 2024. Blackcurrant (fruits, pomace, and leaves) phenolic characterization before and after in vitro digestion, free radical scavenger capacity, and antioxidant effects on iron-mediated lipid peroxidation. Foods 13: 1514. 10.3390/foods13101514
Untea, A.E., Varzaru, I., Panaite, T.D., Gavris, T., Lupu, A., and Ropota, M., 2020. The effects of dietary inclusion of bilberry and walnut leaves in laying hens’ diets on the antioxidant properties of eggs. Animals 10: 191. 10.3390/ani10020191
Untea, A.E., Varzaru, I., Saracila, M., Panaite, T.D., Oancea, A.G., et al. 2023. Antioxidant properties of cranberry leaves and walnut meal and their effect on nutritional quality and oxidative stability of broiler breast meat. Antioxidants 12: 1084. 10.3390/antiox12051084
Varzaru, I., Oancea, A.G., Vlaicu, P.A., Saracila, M., and Untea, A.E., 2023. Exploring the antioxidant potential of blackberry and raspberry leaves: Phytochemical analysis, scavenging activity, and in vitro polyphenol bioaccessibility. Antioxidants 12: 2125. 10.3390/antiox12122125
Varzaru, I., Untea, A.E., Panaite, T., and Olteanu, M., 2021. Effect of dietary phytochemicals from tomato peels and rosehip meal on the lipid peroxidation of eggs from laying hens. Archives of Animal Nutrition 75: 18–30. 10.1080/1745039X.2020.1813515
Vlaicu, P.A., Oancea, A.G., Saracila, M., Varzaru, I., and Untea, A.E., 2025. Unveiling the nutrients profile of wild black elderberry (Sambucus nigra L.) flowers, berries and leaves from Northern and Southern Romanian regions as potential source of pro-health bioactives in human nutrition. Applied Food Research 5: 101139. 10.1016/j.afres.2025.101139
Vlaicu, P.A., and Untea, A.E., 2024. Application of natural antioxidants from fruits waste for improving egg quality characteristics. Applied Sciences 14: 10437. 10.3390/app142210437
Vlaicu, P.A., Untea, A.E., Lefter, N.A., Oancea, A.G., Saracila, M., and Varzaru, I., 2024. Influence of rosehip (Rosa canina L.) leaves as feed additive during first stage of laying hens on performances and egg quality characteristics. Poultry Science 103: 103990. 10.1016/j.psj.2024.103990
Vlaicu, P.A., Untea, A.E., Turcu, R.P., Panaite, T.D., and Saracila, M., 2022. Rosehip (Rosa canina L.) Meal as a natural antioxidant on lipid and protein quality and shelf-life of polyunsaturated fatty acids enriched eggs. Antioxidants 11: 1948. 10.3390/antiox11101948
Yadav, A.S., 2023. Collating antioxidant, reducing and metal chelating properties of spices and Acacia. Food Chemistry Advances 2: 100257. 10.1016/j.focha.2023.100257
Yuan, J., Roshdy, A.R., Guo, Y., Wang, Y., and Guo, S., 2014. Effect of dietary vitamin A on reproductive performance and immune response of broiler breeders. PLoS One 9(8): e105677. 10.1371/journal.pone.0105677
Zhao, H., Chen, Y., Wang, S., Wen, C., and Zhou, Y., 2021. Effects of dietary natural vitamin E supplementation on laying performance, egg quality, serum biochemical indices, tocopherol deposition and antioxidant capacity of laying hens. Italian Journal of Animal Sciences 20: 2254–2262. 10.1080/1828051X.2021.2002733